{kind=link}
垄作模式下甜高粱对农田渍害胁迫的响应
[同琳静1, 2  , 冯涛
, 冯涛1 , 孙政国1 , 陈小东3 , 杨文祥4 , 李建龙1, 2 ]
, 冯涛, 陈小东]
|
|


第一作者:同琳静(1995-),女,陕西渭南人,在读硕士生,主要从事草业科学、农业遥感等研究。E-mail:TLJ1654@163.com
我国长江中下游地区常年降水较多,易形成农田渍害。为探究渍害胁迫对甜高粱( Sorghum dochna)产量性状和抗氧化酶系统的影响,设计了平作和垄作两个处理,在甜高粱三叶期对其进行渍水处理,测定并比较各处理的抗氧化酶系统活性和产量性状。结果表明,苗期遭受渍害胁迫,垄作处理下甜高粱的茎粗、单株干重、叶数、叶面积指数均显著高于平作( P<0.05)。株高和茎叶比在两个处理间无显著差异( P>0.05);与平作处理相比,渍害胁迫下甜高粱叶片的抗氧化酶[超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)]活性更强。研究表明,垄作耕作方式能够提高甜高粱耐受渍害胁迫的能力,获得更高的产量。
Waterlogging is easily caused by common and heavy precipitation in the middle and lower reach regions of the Yangtze River in China. In this study, two culture patterns, ridge and flat, were conducted to determine the effect of culture patterns on the yield traits and antioxidant enzyme activity of sweet sorghum. The yield traits and antioxidant enzymes activity were evaluated and compared after applying waterlogging stress at the seedling stage. The results showed that when the sweet sorghum suffered waterlogging stress at the seedling stage, the stem diameter, dry weight per plant, number of leaves, and leaf area index (LAI) under the ridge culture pattern were significantly higher than those under the flat culture pattern ( P<0.05). Meanwhile, the height and stem and leaf ratio of plants under ridge culture showed little differences compared to those under the flat culture pattern ( P>0.05), and waterlogging stress had less impact on the antioxidant enzyme activities; SOD, POD and CAT were continually highly expressed under the ridge culture pattern. In conclusion, the experiment demonstrated that the application of ridge pattern culture improved the adaptability and yield of sweet sorghum under waterlogging stress.
渍灾是指土壤排水不畅或地下水位过高引起土壤根系缺氧、肥力失调, 从而影响作物正常生长发育的现象[1]。我国每年发生渍害的面积达144.4万hm2, 主要分布在四川、重庆、湖北、湖南、安徽等地, 粮食生产损失严重[2, 3]。
甜高粱(Sorghum dochna)属于一年生禾本科植物, 抗逆性强、适应性广、生物量大。不仅是优质的粮食作物, 同时已成为许多国家优质饲草的重要来源, 此外, 甜高粱还可用作能源作物和糖料作物。国内外学者对于甜高粱的选育、栽培和利用等研究较多[4]。逆境胁迫研究方面以盐胁迫和干旱胁迫居多, 对于渍害胁迫下植物的生理变化和易发渍害胁迫地区的栽培管理技术报道较少。
我国长江中下游地区, 每年的6月下旬至7月上旬会进入梅雨季节, 长时间降水对甜高粱的产量性状及抗氧化酶系统造成影响, 从而降低了甜高粱的产量。研究表明, 垄作耕作方式相比于平作能增加土壤透气性、降低田间湿度, 提高作物产量[5]。为此, 针对江苏里下河地区高湿多雨的气候特点, 设置了垄作和平作两种处理, 探究渍害胁迫下, 垄作和平作两种方式种植甜高粱的产量性状和生理特性的变化规律, 以期为易产生渍害胁迫地区甜高粱种植提供理论依据。
试验所用材料为甜高粱(大力士), 由百绿(北京)国际草业有限公司提供。
1.2.1 试验区概况 试验于2016年在江苏省兴化市华夏畜牧有限公司进行。该地位于长江中下游地区, 属于北亚热带湿润性季风气候, 年平均气温15.0 ℃, 年平均降水量1 032.3 mm, 年平均降水日(日降水量≥ 0.1 mm)109 d。兴化市每年于6月20日左右入梅, 7月10日左右出梅, 梅雨期降水量平均为240 mm; 土质疏松, 耕作层深厚, 肥力中等, 适合甜高粱的生长; 前茬作物为小黑麦(× Triticale Wittmack)。试验地块土壤有机质含量为20.63 g· kg-1, 碱解氮含量为131 mg· kg-1, 有效磷含量为55.47 mg· kg-1, 有效钾含量为78.31 mg· kg-1。
1.2.2 试验设计 采用单因素随机区组设计, 小区面积为3 m× 3 m, 设置了垄作和平作两个处理, 3个重复, 小区设置大沟30 cm, 小沟20 cm。垄作每小区起3垄, 相邻垄之间间距为30 cm, 垄高15 cm, 垄面宽50 cm, 垄上种两行, 行距30 cm。平作设计为宽窄行, 宽行为80 cm, 窄行为30 cm。单株株距为10 cm, 播种量为2.001× 105株· hm-2。播种前施入基肥(复合肥225 kg· hm-2, N:P:K=20:8:18)。
2016年6月15日进行播种, 苗期施入尿素75 kg· hm-2。在三叶期进行间苗、定苗, 并对小区进行渍水处理, 水量保持在排水沟的3/4处, 保证田间持水量为100%。并在渍水处理后的0、3、6、9 d(7月11日、7月14日、7月17日、7月20日)取样测定叶片超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性。测定甜高粱的农艺性状时共进行4次取样(每隔20 d左右), 时间分别为7月11日(苗期)、8月3日(拔节期)、8月24日(拔节期)、9月14日(孕穗期), 取样后测定两种耕作方式下甜高粱的干鲜重、株高、茎粗、单株叶面积、叶数等指标。
1.2.3 指标测定
株高:每个小区选取3株长势均匀的植株, 测量拉伸株体长度。茎粗:每个小区选3株长势均匀的植株, 用游标卡尺测定根部向上第2节中间茎的直径, 记作茎粗。单株叶面积:每个小区选3株长势均匀的植株, 茎叶分离, 用叶面积仪测得单株叶面积, 通过相关换算得到甜高粱的叶面积指数。单株鲜重和干重:每个小区选取3株长势均匀的植株, 留茬5 cm刈割, 茎叶分离后分别测量茎、叶鲜重。然后放入105 ℃烘箱中杀青15 min, 在65 ℃下烘干至恒重, 称重后得到茎、叶干重, 通过计算可得到茎叶比。
抗氧化酶系统的活性测定:在7月11日、7月14日、7月17日、7月20日进行取样, 每个小区取样3份。选取甜高粱最上部两片完全展开叶, 经蒸馏水冲洗吸干, 称取约0.3 g鲜样加8 mL PBS(0.05 mol· L-1, pH 7.8)于冰浴中碾磨, 4 ℃下3 000 r· min-1离心20 min, 取上清液用于抗氧化酶活性的测定, 重复测定3次[6]。采用氮蓝四唑法测定SOD活性[7], 愈创木酚显色法测定POD活性[8], 紫外吸收法测定CAT活性[8]。
试验数据统计采用SPSS 20.0软件对两个处理间产量性状和抗氧化酶活性进行独立样本T检验, 采用Microsoft Excel 2007作图。
苗期受到渍害胁迫, 不同处理下甜高粱的株高、茎粗、单株干重、叶面积指数、叶数、茎叶比都表现为不断增加, 但变化的幅度有所差别(表1)。
2.1.1 株高 7月11日和8月3日, 垄作处理的株高分别比平作高9.76和18.37 cm, 存在显著差异(P< 0.05)。8月24日和9月14日, 两种处理间株高无显著差异(P> 0.05)(表1)。
2.1.2 茎粗 垄作处理的茎粗在初期(7月11日和8月3日之间)增加速率大于平作, 之后增加幅度不大。7月11日, 甜高粱的茎粗在两处理间无显著差异(P> 0.05)。在8月3日、8月24日和9月14日, 垄作处理的甜高粱的茎粗分别比平作高3.27、4.79、5.04 mm, 差异显著(P< 0.05)(表1)。
2.1.3 单株干重 垄作处理的单株干重和单株干重的增加速率始终大于平作。7月11日, 垄作处理的甜高粱的单株干重与平作间无显著差异(P> 0.05)。在8月3日、8月24日和9月14日, 垄作处理的单株干重显著大于平作(P< 0.05)。9月14日, 垄作处理的单株干重为70.78 g, 比平作高29.77 g(表1)。
2.1.4 叶面积指数 垄作处理的甜高粱的叶面积指数增加速率始终大于平作。7月11日, 垄作处理的叶面积指数大于平作, 但无显著差异(P> 0.05)。8月3日、8月24日和9月14日, 垄作处理的叶面积指数分别比平作高0.10、0.79、1.08, 差异均显著(P< 0.05)(表1)。
2.1.5 叶数 在7月11日、8月3日和8月24日, 垄作处理的叶数不仅大于平作, 而且增加速率也大于平作。8月24日至9月14日之间, 平作处理的叶数增加速率大于垄作, 但叶数少于垄作。7月11日, 垄作处理下的叶数与平作间无显著差异(P> 0.05)。在8月3日、8月24日和9月14日, 垄作处理的叶数显著大于平作(P< 0.05), 分别比平作多1.02个、3.22个、2.27个(表1)。
| 表1 渍害胁迫下不同耕作方式对甜高粱不同时期产量性状的影响 Table 1 Effect of different culture patterns on yield traits of sweet sorghum under waterlogging stress |
2.1.6 茎叶比 垄作处理茎叶比的增加速率总体大于平作。7月11日, 垄作处理的茎叶比与平作间无显著差异(P> 0.05)。8月3日, 垄作处理的茎叶比比平作高0.15, 差异显著(P< 0.05)。8月24日和9月14日, 垄作处理的茎叶比与平作间无显著差异(P> 0.05)(表1)。
2.2.1 SOD活性 甜高粱苗期遭受渍害胁迫, 平作处理的SOD活性先升高后降低, 处理6 d时达到峰值, 为318.4 U· (g· min)-1, 而垄作耕作方式下的SOD活性不断升高。取样0~3 d, 平作处理的SOD活性和SOD活性升高速率都大于垄作。取样3~6 d, 虽然平作处理的SOD活性仍大于垄作, 但升高速率开始减慢, 此时垄作处理的SOD活性升高速率大于平作。在取样6~9 d, 平作处理的SOD活性已经开始下降, 而垄作处理的SOD活性还在不断升高。在渍水处理9 d后, 垄作和平作处理的SOD活性分别为304.2和217.2 U· (g· min)-1(图1)。
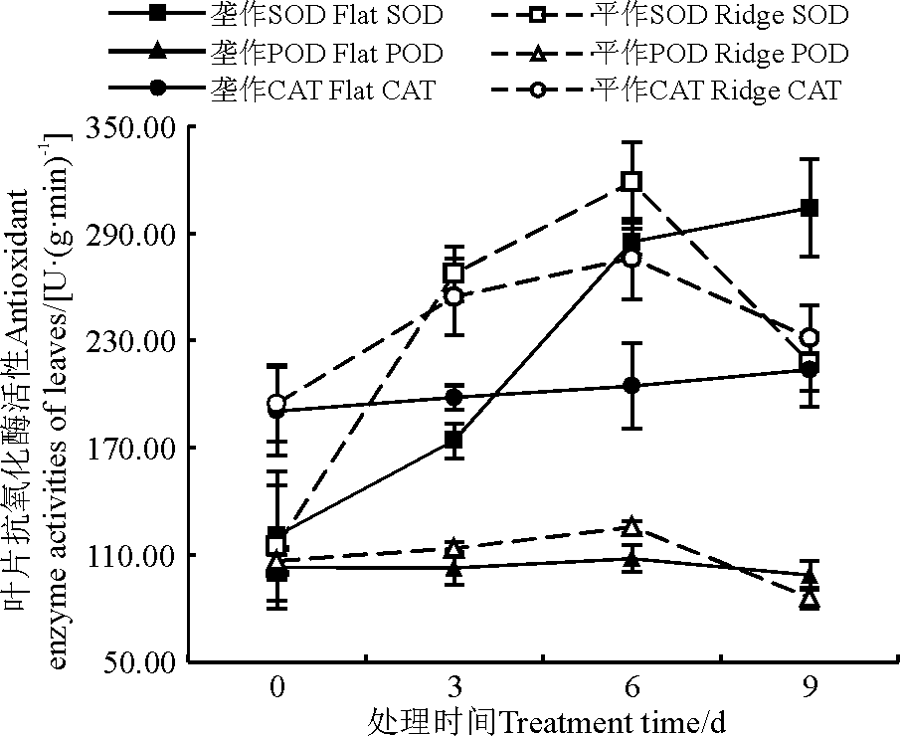 | 图1 渍害胁迫下不同耕作方式甜高粱叶片抗氧化酶活性的变化Fig. 1 Variation trend in the antioxidant enzyme activities of sweet sorghum leaves in two culture patterns under waterlogging stress |
2.2.2 POD活性 苗期渍害胁迫下, 两种处理的甜高粱的POD活性都表现为先升高后降低, 胁迫处理6 d时, 垄作和平作处理的POD峰值分别为108和125.5 U· (g· min)-1。取样0~3 d和3~6 d, 平作处理的POD活性和POD活性升高速率都大于垄作。在取样6~9 d, 两种处理的POD活性都表现为不断降低, 平作处理的降低幅度大于垄作。渍害9 d后, 垄作处理的POD活性比平作高12.8 U· (g· min)-1(图1)。
2.2.3 CAT活性 平作处理的CAT活性先升后降, 在胁迫处理6 d时达到峰值, 为275.8 U· (g· min)-1。而垄作处理的CAT活性不断升高, 取样间隔内的CAT升高速率变化不大。胁迫9 d内, 平作处理的甜高粱的CAT活性始终高于垄作, 但并不能表明平作方式能够更好地表达CAT酶来抵御渍害胁迫。渍害胁迫6~9 d, 虽然平作处理的CAT活性高于垄作, 但其CAT活性表现为降低趋势, 而垄作处理的CAT活性在不断升高, 其活性有超过平作的趋势(图1)。
逆境胁迫对于植物生长发育的影响最终体现在农艺性状上。渍害对玉米(Zea mays)[9]、芝麻(Sesamum indicum) [10]、小麦(Triticum aestivum)[11]、大豆(Glycine max)[12]和棉花(Gossypium hirsutum)[13]等的生长都产生影响, 产量出现降低。玉米在苗期遭受渍害胁迫后, 根的形态发生了变化, 根的条数、长度、干重、吸收营养的能力都显著降低[14]。分别在芝麻的蕾期、盛花期和终花期对其进行相同时间的渍害胁迫, 结果均表现为根系早衰甚至烂根、蒴果数减少、种子产量降低[15]。当甜高粱受到逆境胁迫时, 同样会出现有氧呼吸减弱, 无氧呼吸作用增强, 所以糖类消耗, 酒精大量积累, 根系活动受阻, 代谢缓慢[16]。渍涝环境会使得土壤中的空气含量降低, 溶解氧减少, 无氧呼吸增强, 从而抑制根的正常生理代谢, 影响其对营养物质的吸收。同时, 也会使根的形态发生变化, 改变光合产物在地上部和地下部的分配, 增加植物的冠根比[14, 17]。植物的地上部分和地下部分是相互作用的, 根系活动受阻会导致地上部分的叶绿素合成受阻、蒸腾作用和呼吸作用的速率降低、蛋白质合成减少、氧化磷酸化过程受到抑制, 激素分泌过程也出现紊乱。地上部分生长受到减缓, 同时也会抑制其根的代谢, 从而使植物进入一个死循环, 严重时会导致死亡[18, 19]。所以, 缓解渍害对植物生长的影响要从缓解植物根系的胁迫入手。相关研究[2]表明, 垄作耕作方式能够使作物增加保苗数、叶片的叶绿素含量、干物质积累和叶面积指数, 提高光能利用率、增加土壤透气性、降低田间湿度, 获得了更好的产量性状。赵贵元等[20]发现, 在盛花期对芝麻(Sesamum indicum)进行渍害处理, 垄作方式下的单株产量、株高、有效穗长与平作间存在极显著差异, 能有效缓解渍害胁迫对植物产量性状的影响。本研究结果与在芝麻上的研究结果相似, 垄作处理下甜高粱的茎粗、叶数、叶面积指数都显著大于平作(P< 0.05), 株高和茎叶比等表现出垄作高于平作的趋势。说明垄作耕作方式可缓解渍害胁迫对于甜高粱生理过程的抑制作用, 从而表现出更好的产量性状。
甜高粱苗期受到渍害胁迫下表现出来的抗氧化能力, 是机体发生多种生化反应, 产生多种酶协同作用的结果。SOD、POD、CAT是植物体内重要的3种抗氧化酶, 其功能为清除植物机体内的自由基, 使机体免受伤害[21]。SOD是活性氧清除反应中第一个发挥作用的抗氧化酶, 可以有效防御活性氧对于植物的毒害[22]。一般认为, 植物在遭受各种逆境胁迫(干旱、盐害、重金属等)时, 会增加SOD活性以抵御逆境伤害。但遭受长时间的逆境胁迫时, 自由基和活性氧的增加会超过植物体的调控范围, 从而显著降低SOD活性[23]。甜高粱在苗期遭受盐胁迫时, SOD随胁迫时间的变化表现为先增后降[24], 本研究平作处理与该变化趋势相同, 而垄作表现为不同, 呈现不断增加的趋势。在遭受渍害胁迫的初期, 平作和垄作处理都提高SOD活性来抵御胁迫, 其发生机理为缺氧条件造成自由基合成增加会启动缓解损伤细胞的保护机制, 促进植物机体SOD的表达[6]。其中, 平作处理初期SOD活性升高较快, 说明平作处理对于渍害胁迫更加敏感。随着胁迫时间的延长, 因其机体内的自由基和活性氧会出现积累, 不同处理下SOD的调控能力表现不同。在胁迫9 d后, 平作处理的SOD活性出现下降, 垄作处理的SOD活性大于平作, 并且还表现为不断升高, 说明垄作SOD能够持续有效的清除自由基和活性氧, 更好地抵御渍害胁迫。
POD在植物体内分布广泛, 与光合作用、生物氧化等多种代谢过程有关, 可用于消除自由基, 其活性与植物的抗逆性有关。一般在植物体内, SOD和POD协同作用将H2O2转变为H2O, 防止膜系统遭到破坏[25]。随着植物的生长发育阶段和外界环境的不同, POD活性也表现为不同[22]。本研究结果表明, 不同处理下, POD活性的变化趋势与SOD活性在胁迫初期表现的趋势类似, 平作处理的POD活性和POD活性升高速率都大于垄作。但是随着胁迫时间推移, 两种处理的POD活性都开始出现下降, 原因与SOD活性变化原因相同, 两种处理下甜高粱POD活性变化趋势与苗期遭受盐胁迫时相同[24], 与小麦灌浆期遭受渍害, 旗叶POD活性先降后增的趋势不同[26]。在胁迫结束时, 垄作处理的POD活性已经大于平作。说明垄作处理下POD的表达能更好地抵御渍害胁迫。
研究发现, 随着环境胁迫的加重, 植物的CAT活性会出现持续下降[27]。但也有研究报道, CAT活性在胁迫早期活性会表现出升高, 后期出现不断下降[28]。同时, 也可能出现CAT随胁迫进行, 活性升高的现象[29]。本研究结果表明, 平作和垄作处理下的甜高粱都积极表达CAT以应对渍害胁迫, 初期活性都表现为不断升高, 但后期变化趋势不同, 平作处理的CAT的变化趋势为先升后降, 而垄作处理下则不断增加。胁迫进行9 d后, 垄作处理的CAT活性还在不断增加, 说明垄作处理的CAT能够持续表达, 抵御渍害胁迫对植物内在生理的影响。
农作物的产量基本取决于光合系统的大小和效率, 而叶数和叶面积指数是影响光合作用效率的直接因素[30]。株高、茎粗、茎叶比和叶数是反映植物生长状况的重要农艺性状[31]。渍害胁迫下, 植株体内SOD、POD、CAT、硝酸还原酶(NR)、核酮糖-1, 5-二磷酸羧化酶(Rubisco)等保护酶类, 活性都会出现下降[1]。其中SOD、POD、CAT可有效防止膜氧化; NR 参与N代谢过程, 是最重要的限速酶, 其活性强弱影响 N 代谢水平; Rubisco是决定光合作用效率的关键酶[1]。本研究结果显示, 甜高粱苗期SOD和POD活性都与产量性状具有正相关关系, 这与前人研究中得到的抗氧化酶活性与产量性状正相关的结论相同[19]。CAT活性与产量性状间相关关系都不显著, 可能与本研究中抗氧化酶指标测定仅持续到胁迫后9 d, 有关9 d之后两种不同处理下的CAT酶活性与产量的关系还需要进一步探索。结果说明, 垄作处理下, 渍害胁迫对于细胞酶活性和代谢速率影响较小, 并相比于平作光合效率较高、农艺性状较好、干物质积累较多。
综上所述, 垄作的耕作方式能够通过缓解渍害胁迫对于甜高粱根系生理活动和代谢速率的影响, 更好地表达SOD、POD、CAT等酶活性来适应胁迫环境, 从而相对于平作表现出更好的株高、茎粗、单株干重、叶面积指数、叶数、茎叶比等产量性状。因此, 针对长江中下游地区的气候特点, 垄作的耕作方式更有利于甜高粱的优质生产。
(责任编辑 王芳)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|