

{kind=link}
{kind=link}
{kind=link}
{kind=link}
藏北高寒草甸土壤线虫群落结构对增温的响应
[王学霞1, 2  , 高清竹
, 高清竹1 , 干珠扎布1 , 胡国铮1 , 栗文瀚1 ]
, 高清竹, 栗文瀚|
|


第一作者:王学霞(1982-),女,山东沂源人,助研,博士,主要从事草地生态学研究。E-mail:wxx0427@163.com
为揭示增温对高寒草甸土壤线虫群落的影响,利用OTC模拟短期和长期增温对藏北高寒草甸土壤线虫群落进行比较研究。结果表明,短期和长期增温改变了土壤线虫的群落组成,增加了双垫刃属( Ditylenchus)和丽突属( Acrobeles)丰度。长期增温导致食真菌类线虫丰度显著增加,但各处理间食细菌类线虫、植物寄生类线虫、杂食/捕食类线虫丰度以及cp1-5类群的丰度和属数量无显著差异( P>0.05)。短期和长期增温均降低了土壤线虫的多样性和均匀度,其中2015年短期增温处理显著降低了其多样性。2015年和2016年短期和长期增温土壤显著降低了线虫数量,较对照分别降低了34.45%、32.09%和25.34%和22.66%。各处理样地间MI(Maturity index)、NCR(Nematode channel ratio)、PPI(Plant parasite index)和WI(Wasilewska index)指数无显著差异( P>0.05),且均表现出WI>1,NCR>0.5,表明增温对高寒草甸的健康状态影响不大,土壤有机质矿化途径主要由食细菌和真菌线虫参与。环境因素与土壤线虫数量冗余分析表明,植物总盖度、莎草科盖度、土壤温湿度、细菌和真菌数量对土壤线虫数量影响达到显著水平( P<0.05),增温通过改变植物、土壤理化性质和微生物数量等环境因子而影响高寒草甸土壤线虫群落组成。
To understand the effect of warming on soil nematode communities in alpine meadows, soil nematode communities were compared in short-term and long-term warming by OTC (open top chamber)simulation in northern Tibet. The results showed that the community composition of soil nematodes was changed and the abundance of Ditylenchus and Acrobeles were increased by short-term and long-term warming. The abundance of fungivores (Fu) was increased significantly by long-term warming; however, the abundance of bacterivores (Ba), plant-parasites (Pp), predators/omnivores (Om), and cp1-5 groups was not significantly different among the treatments studied. The diversity and evenness of soil nematodes were reduced by both short-term and long-term warming. The diversity of soil nematodes in 2015 was significantly reduced in the short-term warming treatment. As compared with the control, the number of soil nematodes in the short-term and long-term treatments was significantly decreased in 2015 and 2016. The values were decreased by 34.45% and 32.09% in 2015 and 25.34% and 22.66% in 2016, respectively. The effect of warming on MI, NCR, PPI, and WI index were not significant in the study treatments, and WI > 1, NCR > 0.5, showed that warming had a little effect on the health of alpine meadows. Moreover, the decomposition of organic matter depended mainly on Fu and Ba in this study. The correlations among environmental factors and the number soil nematodes were analyzed by redundancy analysis. The results showed that the total plant cover, Cyperaceae cover, soil temperature and moisture, and number of bacteria and fungi were important factors affecting the number of total nematodes, Ba, Fu, Pp, and Om. Therefore, certain changes in nematode community structure and composition have taken place due to the plants, soil, physical, and chemical properties, and the number of microbes were changed by raising temperature in alpine meadows.
近年来, 以气候变暖为主要特征的全球气候变化对陆地生态系统产生的复杂生态效应已成为科学家广泛研究的热点[1, 2]。土壤线虫作为土壤食物网的重要组成部分, 在土壤系统能量流动和物质循环中起着重要作用, 对气候和环境变化敏感, 成为气候变化的指示生物之一[3]。由于土壤环境相对稳定, 土壤线虫对温度变化的敏感性明显高于地上生物, 土壤温度的微弱变化都会对土壤线虫群落结构和组成发生变化[4, 5]。温度通过影响植被组成特征[6]、土壤特征[7]、土壤生物[8]等多种因素而影响土壤线虫的群落结构和组成。通过不同半干旱区草地生态系统开展的一系列的增温控制试验发现, 增温对土壤线虫结构和组成的影响没有统一的定论, 增温降低了温带草原植物寄生性线虫和捕食/杂食性线虫的丰富度[9]而对其他类群影响不大; 增温降低了温带草原土壤线虫密度[10]; 改变了苔原生态系统和温带草原土壤线虫结构和组成[3, 11]; 对内蒙典型草原土壤线虫组成影响不明显[12]; 增加了苔原生态系统土壤线虫的丰富度、多样性和食物网的复杂性[13]。因此, 增温对土壤线虫结构和组成的影响与当地环境密切相关。
近年来, 作为气候变暖的高敏感地区, 藏北地区气温发生了明显的变化。1960年以来, 青藏高原全年平均温度每10年升高0.2 ℃, 并且在2000年之后温度上升趋势更加明显; 据预测, 到2100年, 高原平均温度将增加2.6~5.2 ℃[14]。高寒草甸生态系统适应长期低温环境, 对气候变暖更为敏感。前人关于模拟温度升高对高寒草甸生态系统植物物种组成和种群结构, 生态系统结构与功能具有重要的影响[15, 16, 17], 也影响植物生理特征和生存策略[18, 19], 同时改变土壤理化性质和营养动态[20]。迄今, 有关于高寒草甸生态系统土壤线虫对增温的响应的研究几乎未见报道。本研究以藏北高寒草甸为研究对象, 通过增温模拟控制试验, 研究短期(1、2年)增温和长期增温(5、6年)对高寒草甸土壤线虫群落结构的影响, 明确增温条件下藏北高寒草甸土壤线虫群落变化趋势及其关键影响因素, 以期增加气候变暖对高寒草甸土壤动物影响的认识。
试验区位于藏北地区那曲县那曲镇的农业部农业环境科学观测实验站内(31.44° N, 92.02° E)。该地区年均温度为0.3 ℃(2005-2014年), 年降水量达461.8 mm, 生长季(5-9月)平均气温为8.0 ℃; 年降水量为461.8 mm。草地类型为高寒草甸, 建群种为莎草科植物高山嵩草(Kobresia pygmaea), 其他莎草科植物包括青藏苔草(Carex moorcroftii), 线叶嵩草(Kobresia capillifolia)和矮生嵩草(Kobresia humilis); 禾本科植物包括早熟禾(Poa pratensis)和丝颖针茅(Stipa capillacea); 豆科植物包括藏豆(Stracheya tibetica)和紫花棘豆(Oxytropis subfalcata); 杂类草包括钉柱委陵菜(Potentilla saundersiana)、二裂委陵菜(Potentilla bifurca)、菊叶委陵菜(Potentilla tanacetifolia)以及肉果草(Lancea tiberica)等。土壤类型为高寒草甸土, 质地为砂壤土[18]。
增温试验:采用开顶式生长室(open top chamber, OTC) 模拟增温对藏北高寒草甸影响的相关试验。OTC 以聚碳酸酯为主要原料, 规格:底部直径1.2 m, 顶部直径0.6 m, 高度0.45 m。试验设置对照处理(CK); 短期增温(SW):增温1年(2015年)和2年(2016年); 长期增温(LW):增温5年(2015年)和6年(2016年)。
土壤取样:每年8月初进行采样, 用直径5 cm土钻取0-15 cm的土壤, 每个样地取两钻混匀为一个样品, 用于土壤动物、微生物培养和养分分析。
植物指标测定:在高寒草地植物旺盛生长期(8月初), 每个样方内选取0.5 m× 0.5 m测定草地植被盖度、物种、地上生物量。土壤理化性质测定:在每个试验样地都装有温度(DS18B20)和湿度(EC-5)探头记录土壤温度和土壤湿度, 记录频率为每15 min一次。用电感耦合等离子体光谱仪(Spectro Arcos Eop, Spectro, German)测定土壤总氮含量, 用元素分析仪(Vario EI, Elementar, German)测定有机碳含量。
微生物培养和分析:采用稀释平板法测定土样中细菌、真菌及放线菌的数量, 在恒温恒湿培养箱30 ° C下恒温培养(上海森信, HWS-450)。采用牛肉膏蛋白胨琼脂培养基培养细菌, 培养3~5 d, 检查菌斑数量。利用马丁孟加拉红培养基培养真菌, 采用改良高氏一号培养基培养放线菌, 均培养5~7 d, 检查菌斑数量。
土壤线虫分离和鉴定:取 100 mL土壤用改进的贝曼浅盘法分离法分离, 室温条件下, 分别经24、36和48 h后收集浅盘水, 过筛、冲洗、保存。当分离出的线虫数量小于150条时, 鉴定所有的线虫种类, 当数量多于150条时, 只鉴定150条, 然后统计所鉴定出的线虫数量。参照《中国土壤动物检索图鉴》[21]和《植物线虫分类学》[22]根据线虫的头部形态学特征和取食生境将土壤线虫划分到属。根据划分原则, 将土壤线虫划分为4个营养类群[23]:食细菌类(bacterivores, Ba)、食真菌类(fungivores, Fu)、食植寄生性线虫(plant-parasites, Pp)、捕食类/杂食类(predators/omnivores, Om)。同时按照colonizer-persister(c-p), 将线虫从c-p1到c-p5值分成5个类群[24]。
土壤跳虫和线虫的多度按以下标准划分:个体数量占总捕获量10%以上者为优势类群(+++); 1%~10%者为常见类群(++); 不足1%者为稀有类群(+)。
采用国内外学者广泛采用的Shannon-Weaver指数(H')、Pielou均匀度指数(J)研究土壤线虫的群落多样性。公式如下:
H'=
J=(-∑ PilgPi)/lnS。
式中:Pi为土壤线虫群落第i类群个体数占总个体数的比例; S为土壤线虫群落中所有线虫类群数。
本研究采用MI(Maturity index of free-living nematode)指数、PPI(Plant parasite index)指数、NCR(Nematode channel ratio)指数、WI(Wasilewska index)指数表达各生境土壤线虫群落功能结构特征。上述指数计算公式如下:
自由线虫成熟度指数:MI=∑ [ν (i)× f(i)];
植物寄生线虫指数:PPI=∑ [ν (i)× f'(i)];
线虫通道指数:NCR=NBF/(NBF+NFu);
瓦斯乐斯卡指数:WI=(NBF+NFu)/NPp。
式中:υ (i)是在生态演替中对属于k-选择和r-选择的属分别赋予的c-p值; f(i)是自由生活线虫属在线虫种群中所占的比例; f'(i)是植物寄生性线虫属在线虫种群中所占的比例; NBF为食细菌线虫数量, NFu为食真菌线虫数量, NPp是植物寄生线虫数量。
所有线虫数量的原始数据经过对数转化保证方差齐性检验。分析过程运用 SPSS 22.0 软件。土壤线虫群落数量运用单因素方差分析(One-way analysis of variance), 差异显著性水平为0.05。排序分析用 CANOCO 4.5软件运行, 线虫群落原始数量数据经过对数转换。蒙特卡洛检验排序轴的统计显著性。
2015年和2016年, 在所有样地中, 每100 g干土中土壤线虫的个体密度在380~702条, 平均为508条(图1)。土壤线虫个体密度均表现出CK> LW> SW, 并且CK处理显著高于SW和LW处理(P< 0.05), SW和LW处理间差异不显著(P> 0.05)。
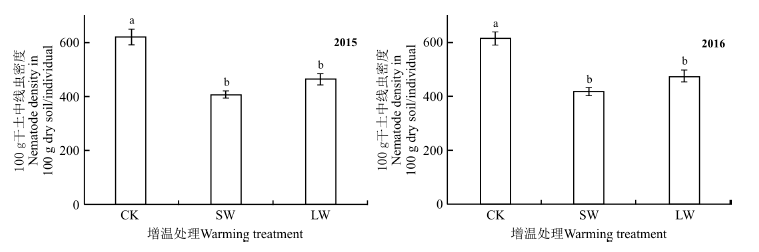 | 图1 增温处理下高寒草甸不同种类土壤线虫的密度 不同小写字母表示表示同一指标不同处理间差异显著(P< 0.05)。CK、SW和LW分别表示对照、短期增温、长期增温。下同。Fig. 1 The density of soil nematode trophic groups under warming experimental treatments in the alpine meadows Different lowercase letters for the same parameter indicate significant differences among different treatments at the 0.05 level. CK, SW and LW indicate control, short-term and long-term warming, respectively; similarly for the following tables and figures. |
2015年和2016年共分离得到土壤线虫2 896条, 49个属, 其中CK处理样地共分离得到土壤线虫1 237条46属, SW处理样地共分离得到土壤线虫619条44属, LW处理样地共分离得到土壤线虫940条44属。其中食细菌线虫18属, 食真菌线虫7属, 植食性线虫14 属, 捕食-杂食线虫10 属(表1)。在所有处理样地中, 食细菌线虫均为优势类, 占个体总数的29.7%~42.1%; 其次是植物寄生类线虫, 占个体总数的28.1%~42.1%; 再次, 杂食/捕食类线虫, 占个体总数的14.9%~20.6%, 食真菌类线虫丰度最低(除LW处理), 占个体总数的12.1%~23.6%。2015年LW处理食真菌类线虫显著高于SW处理(P< 0.05)(图2), 而2015年LW处理显著高于CK(P< 0.05)。
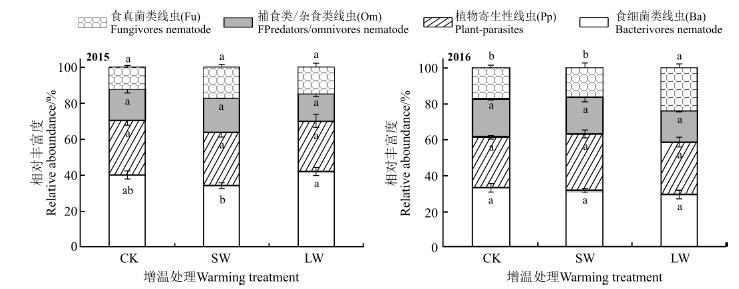 | 图2 增温处理下高寒草甸土壤线虫营养类群的相对丰富度Fig. 2 The relative abundance of soil nematode trophic groups under warming experimental treatments in the alpine meadows |
| 表1 增温处理下高寒草甸土壤线虫群落组成及营养类群及c-p值 Table 1 Compositions of nematode community and trophic groups and c-p values under warming experimental treatments in the alpine meadows |
短期增温和长期增温对土壤线虫(属)组成有一定的影响。在所有处理样地中, 矮化属(Tylenchorhynchus)、中小杆属(Mesorhabditis)和类短体属(Pratylenchoides)均为优势属, 而拱唇属(Labronema)为CK样地的优势属, 双垫刃属(Ditylenchus)为SW和LW样地的优势属, 头叶属(Cephalobus)为LW样地的优势属。拟丽突属(Acrobeloides)等常见属, 真头叶属(Eucephalobus)等稀有属占总属数量的85%以上(表1)。小杆属(Rhabditida)是CK样地特有属, 丽突属(Acrobeles)是LW和SW样地特有属。增温处理没有显著改变高寒草甸线虫功能类群组成(图3)。在获得的所有线虫中, cp2个体和属数量最大, 其次是cp4个体和属数量, 再次是cp3个体和属数量, cp1的属最少, cp5 的个体数量最少。
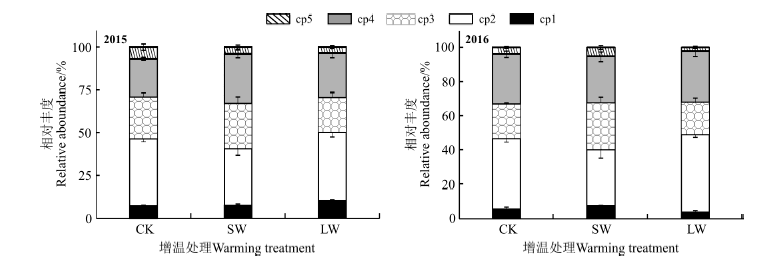 | 图3 增温处理下高寒草甸土壤线虫功能类群丰度(cp) cp1, …cp5表示按照colonizer-persister分成的5个线虫类群。Fig. 3 The abundance of soil nematode functional groups (cp) under warming condition in the alpine meadows cp1, …cp5 indicate 5 groups divided by colonizer-persister. |
采用线虫多样性指数(H’ )、均匀度指数(J)来评价线虫群落的多样性。多样性指数(H’ )表现出CK> LW> SW, 但处理间差异不显著(P> 0.05); 均匀度指数 J与H’ 表现趋势一致, 但是2015年SW处理显著低于CK(P< 0.05), 表明短期增温处理降低了线虫丰度分布均匀和群落结构稳定性, 尤其是短期增温处理(表2)。
自由生活线虫成熟度MI指数, 线虫通路比值NCR指数、植物寄生线虫成熟度PPI指数、瓦斯乐斯卡WI指数表达不同处理土壤线虫群落功能结构特征(表2)。不同处理间线虫MI、PPI指数差异不明显, 表明样地受增温干扰较小, 植地上植被生长状态较好。在所处理样地的WI值> 1, 表明矿化途径主要由食微线虫参与, 土壤处于相对健康状态。短期和长期增温处理对土壤健康状态影响不大。所处理样地的NCR 值都大于0.5, 表明土壤有机质的分解途径均以细菌为主, 因此, 短期和长期增温处理没有改变土壤有机质的分解途径(表2)。
| 表2 增温处理下高寒草甸土壤线虫生态指数 Table 2 Nematode ecological indices under experimental treatments in the alpine meadow |
2015和2016年, 植物生物量均表现为LW> CK> SW, LW处理显著高于CK、SW处理(P< 0.05)。2015和2016年, SW处理植被总盖度和莎草科植物盖度显著低于CK, 而禾本科和杂类草盖度差异不明显。2015年和2016年, 细菌数量和真菌数量均表现出为CK> LW> SW, 2016年SW处理细菌数量显著低于CK(P< 0.05), 放线菌数量表现出相反的趋势。2015和2016年, 土壤温度均表现为SW> LW> CK, 其中SW和LW处理显著高于CK(P< 0.05), 增温幅度分别为0.67、0.73和0.63、0.68 ℃。土壤湿度均表现为CK> LW> SW, CK处理显著高于SW和LW处理(P< 0.05)。不同处理间土壤有机碳和全氮含量均表现为CK> LW> SW, 各处理间差异不显著(P> 0.05)(表3)。
| 表3 增温处理下高寒草甸植物、土壤和微生物指标 Table 3 The plants, soil and microbiological indexes were analyzed under warming experimental treatments in the alpine meadow |
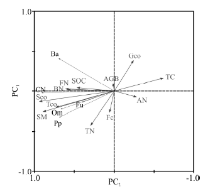
冗余分析表明, 植物、土壤理化性质和微生物数量是导致土壤线虫数量在不同处理间存在差异的重要原因, 12个环境变量解释了77.3%的数据总差异, 其中第一轴和第二轴的解释率为69.2%和4.1%(表4)。食细菌线虫(Ba)数量与食真菌线虫(Fu)数量与细菌数量(BN)、真菌数量(FN)、有机碳(SOC)、土壤湿度(SM)、植物生物量(AGB)、总盖度(Tco)、莎草科盖度 (Sco)、杂草盖度(Fco)呈现正相关; 而与温度(TC)、放线菌(AN)数量呈现负相关。线虫总数(CN)、捕食/杂食类线虫(Om)与细菌数量(BN)、真菌数量(FN)、土壤湿度(SM)、有机碳(SOC)、全氮(TN)、总盖度(Tco)、莎草科盖度(Sco)、杂草盖度(Fco)呈现正相关, 而与温度(TC)、放线菌(AN)数量和禾本科盖度(Gco)呈现显著负相关。植物寄生线虫数量(Pp)与细菌数量(BN)、土壤湿度(SM)、有机碳(SOC)、全氮(TN)、总盖度(Tco)、莎草科盖度(SCover)呈现正相关, 而与温度(TC)和禾本科盖度(Gco)呈负相关关系(图4)。根据箭头的长短可知, 对土壤线虫数量影响程度最大的环境因素是Sco, 最小的是AGB。蒙特卡罗检验结果表明, Sco(F=31.4, P< 0.002), SM(F=24.9, P< 0.002), Tco(F=12.1, P< 0.01), FN(F=7.3, P< 0.02), TC(F=7.2, P< 0.02), BN(F=6.3, P< 0.02)对土壤线虫数量影响达到显著水平。因此, 莎草科盖度、土壤温湿度、植物总盖度、土壤细菌数量、真菌数量是影响高寒草甸土壤线虫数量关键环境因子。增温通过改变植物特征、土壤理化性质和微生物数量等环境因子而影响高寒草甸土壤线虫群落组成。
| 表4 基于线虫物种数量与环境因子的RDA分析 Table 4 Redundancy analysis results for environmental factors and soil nematode number in the alpine meadows |
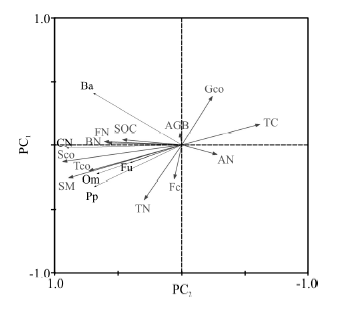 | 图4 高寒草甸土壤线虫数量与环境因子之间RDA分析 AGB, Tco, Sco, Fco, Gco, BN, FN, AN, TC, SM, SOC, TN分别表示地上生物量、总盖度、莎草科盖度、禾本科盖度、杂类草盖度、细菌数量、真菌、放线菌、土壤温度、土壤湿度、有机碳、全氮。Ba, Fu, Pp, Om, CN分别表示食细菌类、食真菌类、植物寄生线虫、捕食类/杂食类, 线虫总数。Fig. 4 The RDA ordination digram of soil nematode number and environmental factors in the alpine meadows AGB, Tco, Sco, Fco, Gco, BN, FN, AN, TC, SM, SOC and TN indicate aboveground biomass, total cover, Cyperaceae cover, Gramineae cover, forb cover, bacteria number, fungi number, Actinomyces number, soil temperature, soil moisture, soil organic carbon, total nitrogen and The total number of nematode, respectively. Ba, Fu, Pp, Om, CN indicate the number of bacterivores, fungivores, plant-parasites, predators/omnivores and the total number of nematode. |
本研究中植物寄生性线虫和食细菌类线虫是该地区高寒草甸的主要营养类群, 杂食/捕食类线虫和食真菌类线虫的比例较低, 土壤线虫cp2类群均为优势类群, 生活策略以r对策为主, 这与薛会英等[25]在该地区的研究结果一致。研究中长期增温(5、6年)显著增加了群落中食真菌类线虫丰度, 这与Briones等[26]温带草地生态系统得出的结论一致, 这表明可能在持续增温条件下, 高寒草甸系统中可能表现出真菌驱动的食物网增强的趋势。增温对食细菌、植物寄生性、杂食/捕食类群线虫影响不明显。食细菌线虫相对多度可能反映土壤中细菌数量及土壤有机质情况[8, 27], 杂食/捕食性线虫是决定土壤食物网复杂性的关键生物类群, 这说明增温对高寒草甸土壤有机质含量和土壤食物网的复杂性影响不大。
在属的水平上, 短期和长期增温降低了拱唇属(Labronema)丰度, 增加了双垫刃属(Ditylenchus)丰度, 长期增温增加了头叶属(Cephalobus), 与Ruess等[11]和Bakonyi 等[3]增温改变了苔原生态系统和温带草原土壤线虫结构和组成结果不同。增温增加了丽突属(Acrobeles)丰度, 这与Bakonyi 等[3]的结果一致, 丽突属线虫与温度呈正相关关系。增温降低了小杆属(Rhabditida)丰度, 这与Papatheodorou等[28]的结果一致, 小杆属线虫与温度呈负相关关系。因此, 高寒草甸不同属线虫对温度变化的响应不同, 短期和长期增温改变了部分线虫的群落组成。
本研究表明, 长期和短期增温均导致高寒草甸土壤线虫密度均降低, 这与前人在草原生态系统中的研究结果一致[10]。长期和短期增温均降低了线虫群落的多样性, 因此增温处理降低了线虫均匀性和群落结构稳定性, 说明增温对高寒草甸生态系统造成了负面影响。这与Bakonyi和Nagy[29]在草地系统中的得出的结论一致, 与Nielsen等[13]在苔原生态系统得出的结论相反, 这可能是由于不同生态系统中环境因子和增温幅度差异而导致的。
短期和长期增温对MI、PPI、WI线虫功能指数的影响不明显, 表明在增温幅度0.63~0.73 ° C下, 高寒草甸系统植被、土壤处于相对健康状态。在该研究中增温降低了NCR指数, 但是NCR指数> 0.5, 表明高寒草甸有机质分解均主要依靠细菌分解途径与薛会英等[25]结果一致, 在所有处理样地的WI值> 1, 表明矿化途径主要由食微线虫参与, 增温没有改变这种分解途径[30]。
土壤线虫群落动态主要受资源有效性及所处生境微环境变化的调控。前人的研究表明土壤温度、土壤水量[31], 土壤SOC、氮等土壤因子[3, 32]、植物群落组成与生产力[6, 33, 34], 土壤微生物数量种类[8, 27] 等都受环境温度的影响, 是线虫群落结构变化的重要驱动因子[35]。本研究中, 通过RDA分析发现增温对高寒草甸土壤线虫群落结构组成的影响, 与增温改变了高寒草甸植物盖度、生物量、土壤温湿度、SOC、TN以及可培养真菌和细菌的数量关系密切。植物生物量与温度之间呈现负相关关系[34], 植物盖度、生物量与土壤线虫密度呈正比关系[35], 由此, 增温降低了群落植物盖度及生物量, 不能为土壤线虫提供了更多的食物来源和庇护所, 因此降低了土壤线虫的个体密度和多样性。在适合的湿度范围内, 草地土壤线虫种群密度与土壤湿度呈显著的正相关关系[3, 36], 由于土壤线虫本身没有自主运动能力, 其活动依赖于土壤孔隙水进行, 土壤含水量高, 利于土壤线虫的运动及捕食, 扩大其群落规模, 但是OTC增温使得高寒草甸土壤蒸发量增加导致土壤湿度降低, 因此降低了土壤线虫的种群密度。食细菌线虫主要以细菌为食, 增温降低了土壤细菌数量, 因此导致食细菌线虫的丰度有所降低。增温对土壤真菌数量影响不显著[37], 这可能是导致短期增温对真菌为食的线虫丰度影响不明显的主要原因, 但是长期增温增加了食真菌类线虫丰度, 这可能是由于长期增温改变了土壤真菌的群落组成[38], 食真菌类线虫丰度不仅与真菌数量有关还与其组成密切相关。由于短期处理土壤含水量、植物盖度、生物量、SOC、TN以及细菌和真菌数量低于长期增温处理, 由此短期增温土壤线虫密度和多样性低于长期增温。这说明, 随着增温时间增加, 高寒草甸土壤线虫对增温的适应性可能是逐渐增强。
食细菌类线虫、植物寄生性线虫和cp2类线虫是各处理样地土壤线虫的优势营养类群, 短期和长期增温显著降低了土壤线虫密度, 长期增温显著增加了食真菌类线虫丰度。因此, 对照、短期和长期增温处理藏北高寒草甸土壤线虫密度、群落组成均具有一定的差异性, 这是由于增温改变了高寒草甸土壤温湿度、有机质含量、土壤细菌和真菌数量以及植物总盖度和莎草科盖度。短期增温和增温降低了土壤线虫的J指数, 在一定程度上降低了土壤线虫的均匀性和结构的稳定性。增温对土壤线虫H’ 、MI、NCR、PPI、WI指数没有显著差异, 表明增温对高寒草甸的健康影响不大, 土壤呈相对健康的状态。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|