

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
玛曲高寒草甸放牧强度的遥感监测
[马青青1, 2  , 柴林荣
, 柴林荣1, 2 , 马海玲3 , 常生华1, 2 , 程云湘1, 2 ]
, 柴林荣]
|
|


第一作者:马青青(1992-),女,甘肃临夏人,在读硕士生,研究方向为草地监测与评价。E-mail:maqq16@lzu.edu.cn
为给玛曲县高寒草甸草-畜平衡和草地资源的可持续利用提供科学依据,本研究利用遥感技术,结合2016年地面实测数据和同时期的MODIS影像资料,分别建立了研究区草地地上生物量和植被指数(NDVI)、相对放牧强度间的回归模型以及不同相对放牧强度和NDVI值之间的幂回归模型。结果表明,6-9月调查点NDVI和地上生物量间存在较好的相关性,即随着NDVI值的增加生物量呈上升趋势,且9月份的拟合( R2=0.510 2) 达到了极显著水平;实测地上生物量和相对放牧强度之间有很好的拟合关系( R2=0.965 1),随着放牧强度的增加地上生物量呈下降趋势;相对放牧强度和植被指数间相关性较好( R2=0.631),即随着NDVI值的变小,相对放牧强度逐渐增强,当相对放牧强度增强到一定程度时,NDVI对其响应不灵敏。
To provide a scientific basis for the sustainable utilization of grassland resources and grass-livestock balance in the Maqu alpine meadow, this study used remote sensing technology combined with field-measured data and MODIS image data in the same period in 2016. Regression models of aboveground biomass and vegetation index (NDVI) with relative grazing intensity were established individually in the study area, and a power regression model between different relative grazing intensities and NDVI was additionally established. The result showed that with increase in NDVI, aboveground biomass increased in June, July, August, and September. Ground biomass and relative grazing intensity showed optimum significant negative correlation ( R2=0.965 1), which was a power curvilinear regression. Relative grazing intensity and vegetation index showed good correlation ( R2=0.631); with decrease in NDVI, relative grazing intensity increased gradually. When relative grazing intensity increased to a certain extent, NDVI was not sensitive to its response.
青藏高原生态系统对全球气候变化较为敏感[1], 高原生态系统的相关研究在全球变化研究中占据极为重要的地位[2]。高寒草甸生态系统是青藏高原最主要的生态系统类型, 约占青藏高原总面积的35%, 在高原草地畜牧业中占有十分重要的地位[3]。放牧活动是高寒草甸的主要利用方式之一。多数研究表明, 适度的放牧可以有效促进草地生产力的提高和生物多样性的维持, 但过度放牧往往会引起草地的大面积退化以及草地初级生产力和土壤理化性状的恶化[4]。近年来, 由于长期受超载过牧等因素的影响, 高寒草甸生态系统退化显著[5], 从20世纪90年代开始, 尤其青藏高原东缘草地出现了明显的退化, 其基本特征为区域气候趋于干旱、地表径流量显著减少、水土流失日益严重[6], 草畜矛盾日益尖锐。
草地生物量是评价青藏高原生态脆弱性和敏感性的重要指标及度量草地退化的重要指标[7], 也是整个草地生态系统运行的能量基础和物质来源, 不仅是体现草地生态系统稳定性和种群多样性的指标, 同时也被用于指导畜牧业生产[8]。对生物量的定量研究, 可以及时准确地获取草地信息, 为科学合理确定载畜量和有效进行草地资源保护提供参考依据, 同时也为草地生态环境研究提供基础数据。草地生物量是草地资源遥感研究的主要指标之一, 监测草地生物量的时空变化动态能够较好地反映草地的生长状况, 在除放牧外无其他干扰因素的情况下能够指示放牧强度的大小, 对牧区草地退化评价和草畜平衡决策研究具有重要意义[9]。
遥感数据已被广泛应用于大尺度植被活动的研究[10, 11, 12], 其在实现草地估产、确定草地载畜量以及加强草地科学管理等方面都有重要的指导意义[13]。Paruelo等[14]在美国中部草原地区将NOAA/AVHRR遥感数据用于草地资源动态监测, 研究表明, 归一化植被指数与草地地上生物量高度相关, 且发现幂函数回归模型是该区的最佳遥感反演模型。Bé nié 等[15]将遥感数据和社会经济因素加到模型中, 以至少80%的准确性模拟了萨赫勒半干旱地区十多年的生物量, 因其适应性强的模拟能力减少了数据采集成本, 且有助于萨赫勒环境的管理。韩波[16]结合三江源区高寒草地地上生物量(aboveground biomass, AGB)野外实测数据和同期MODIS-NDVI(normalized difference vegetation index)和MODIS-EVI(enhanced vegetation index)数据, 通过回归分析, 建立研究区高寒草地AGB遥感反演模型, 并对三江源区近十年高寒草地AGB时空格局进行模拟分析。周鑫等[17]分析了EVI和NDVI与实测草地地上生物量的一元线性、指数和二次多项式回归模型, 并对各种回归模型进行分析比较, 利用优选模型的反演结果分析了伊犁地区草地地上生物量的空间分布。综合上述研究, 采用MODIS植被指数能够监测和预测草地产量, 只是不同的地区会因实际情况采用不同的指数或模型。但是, 至今为止, 很少有研究直接涉及放牧强度和植被指数之间的关系。张艳楠等[18]利用NDVI和生物量建立的生物量估产模型计算出34家牧户的7、8月生物量的差值, 以放牧强度为应变量, 差值为自变量, 通过回归分析建立了放牧强度估测模型(R2 = 0.461), 发现二者之间具有较好的相关性。该研究中还是以生物量作为桥梁, 若是直接利用NDVI和放牧强度来建立大范围放牧强度估测模型, 一方面突破了放牧强度量化的区域限制, 另一方面可以对放牧行为实时监控, 调整家畜活动范围, 从一定程度上缓解草地退化压力。
本研究首先利用野外62个草地调查样点实测地上生物量数据, 建立了研究区植被指数与草地地上生物量的回归模型, 并结合实测地上生物量和不同相对放牧强度间的相关性, 分析了不同放牧强度条件下草地生物量的空间分布特征。然后利用3家牧户的相对放牧强度, 结合同期MODIS影像获取的植被指数数据, 将相对放牧强度和对应的NDVI在区域空间尺度上建立回归模型, 获取研究区植被景观格局和家畜放牧利用强度的时空分布关系, 以期为研究区草地资源的可持续利用和草畜平衡提供依据。
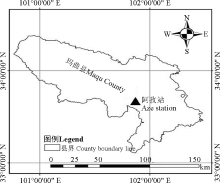
研究区设在甘肃省玛曲县阿孜畜牧科技示范园区(图1), 地理位置为33° 40' N, 101° 52' E, 海拔3 650 m, 年均气温1.2 ℃, 年均降水量620 mm, 主要集中在5-9月, 年日照时数约2 580 h, 年平均霜日大于270 d, 无绝对无霜期, 只有冷暖季之分。土壤为亚高山草甸土。草地类型为高寒草甸, 植被以嵩草(Kobresia sp.)、苔草(Carex sp.)、针茅(Stipa sp.)、羊茅(Festuca sp.)、披碱草(Elymus sp.)和早熟禾(Poa sp.)等为主。牧草一般在4月中、下旬开始返青, 8月中、下旬开始枯黄。放牧家畜主要是地方藏系牦牛。
 | 图1 玛曲县阿孜站的地理位置Fig. 1 Location of Aze station in Maqu County |
试验样地主要设置在阿孜畜牧科技示范园区的牧户地, 选择草地类型一致(高寒草甸为主)、生产和利用方式相似(以牦牛放牧为主)的3家牧户的草地作为研究样地, 而且这些草地都处于同一生长水平(即草地的理论可食牧草产量相等, 且生长周期一致, 除放牧外无其他干扰因素)。根据牧户牦牛数量、草地面积、距离牧户棚圈的远近及牦牛选择摄食的活动路线和在空间停留频次时所表现出的放牧活动强度, 将牧户草地划分为3个放牧梯度区, 即距离畜圈入口较近区域为第一放牧梯度区, 距离畜圈入口中等距离区域为第二放牧梯度区, 距离畜圈入口较远区域为第三放牧梯度区(图2)。在每个放牧梯度区随机选择10个0.5 m× 0.5 m的样方, 且采样点间距符合遥感数据空间分辨率的要求, 每个放牧梯度区重复4次, 在6、7、8、9月的下旬齐地刈割, 进行草地植被调查。记录样方内主要物种的名称、盖度、高度等植物群落信息, 然后齐地面刈割, 称其鲜重, 再在70 ℃烘至恒重称其干重。
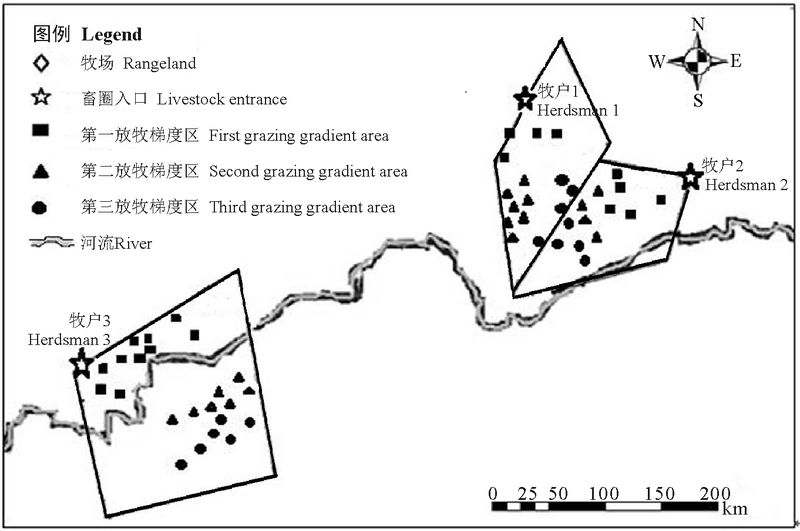 | 图2 试验样地Fig. 2 Sample plot |
归一化植被指数(NDVI)通常是红波段(R)和近红外波段(NIR)反射率之差与之和的比值, 用以表征地表植被的数量分配和质量情况, 即:
NDVI=(NIR-R)/(NIR+R)。 (1)
为了保持与地面实测数据的时空一致性, 选取了2016年6月25日、7月27日、8月28日和9月29日的美国航空航天局(NASA)数据中心提供的16 d合成的250 m空间分辨率的MOD13Q1产品用于提取NDVI值。
根据设定的不同放牧梯度, 计算3家牧户的相对放牧强度:
相对放牧强度=每个牧户家牦牛头数/不同放牧梯度区域面积(不同半径的同心圆面积)。 (2)
相对放牧强度不表示量化的放牧强度, 而是对放牧强度强弱程度的体现。在本研究中, 假定每个放牧梯度的区域是不同半径的圆形, 3个梯度的区域就是3个以牧场入口为圆心的同心圆, 不同梯度上的区域面积就是圆的面积, 且假设同一牧户每个梯度上的牦牛头数相同。虽然与传统放牧强度的算法一样, 即家畜头数除以草地面积, 但因假设每个梯度上牦牛头数相同, 发生变化的只是面积, 故将计算结果定为相对放牧强度, 定性表示放牧的强弱程度。
利用MODIS数据重投影工具软件(MRT)将下载的影像进行NDVI提取, 利用ArcMap软件提取各样方位置点对应的NDVI值, 并与实测生物量建立模型。在野外样点实测过程中, 由于GPS定点的误差以及在称量、烘干过程中出现失误(样品外溢或吸湿), 所以在建立回归模型前需剔除异常值。采用Microsoft Excel和SPSS软件进行相关性的统计分析。
根据公式(2)计算相对放牧强度, 如表1所列。
对比3家牧户间的相对放牧强度, 可以看出, 牧户1的放牧强度比牧户2和牧户3都要大, 牧户3次之, 牧户2则最轻。在同一个放牧梯度水平上, 除了第三个放牧梯度上牧户2和牧户3相对放牧强度相同外, 其他两个梯度上都是牧户1相对放牧强度最大, 牧户3次之, 牧户2最小。
| 表1 3家调查牧户的相对放牧强度 Table 1 Relative grazing intensity of the three herds under survey |
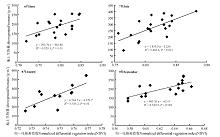
建立调查点植被指数和地上生物量的线性回归模型, 发现6-9月植被指数和地上生物量间的拟合呈相同趋势, 即随着NDVI值的增长生物量呈上升趋势。其中, 9月的拟合效果最好(图3)。
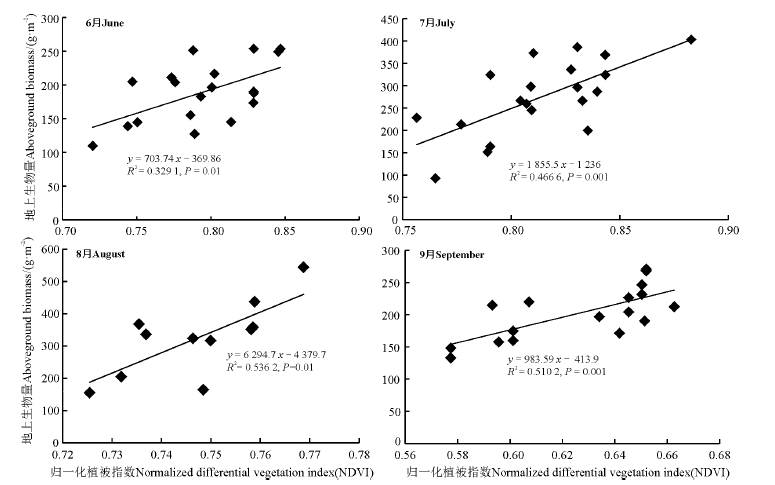 | 图3 调查点植被指数和地上生物量线性回归散点图(6-9月)Fig. 3 Linear regression scatter plots of vegetation index at survey points and aboveground biomass (Jun. to Sep.) |
为进一步研究实测地上生物量和相对放牧强度之间的关系, 建立了6-9月实测地上生物量和相对放牧强度间的回归模型(图4), 发现二者拟合很好(R2=0.965 1), 在某一放牧强度下, 地上生物量达到了最大值, 而后随着放牧强度的增大, 地上生物量呈下降趋势。
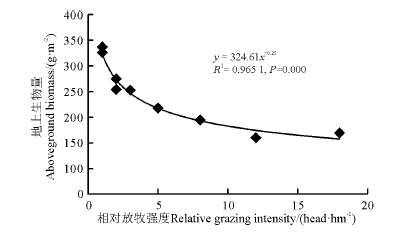 | 图4 实测地上生物量和相对放牧强度的幂回归模型Fig. 4 Power regression model for measured aboveground biomass and relative grazing intensity |
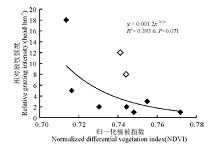
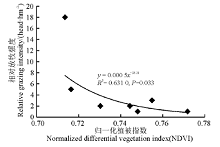
分析了地上生物量和植被指数、相对放牧强度间的相关关系后, 又建立了实验月相对放牧强度与对应梯度上的NDVI值之间的幂回归模型(图5), 结果表明两者之间不存在相关性。在剔除了两个特殊点之后, 得到新的模型(图6), 发现二者相关性较好(R2=0.631 0)。可以看出, 随着NDVI的变小, 相对放牧强度不断增大, 当相对放牧强度达到一定程度后, NDVI对相对放牧强度的响应不再灵敏。
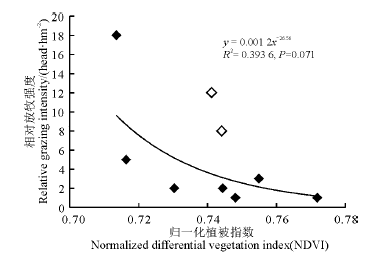 | 图5 调查点植被指数和相对放牧强度的幂回归模型Fig. 5 Power regression model for vegetation index and relative grazing intensity |
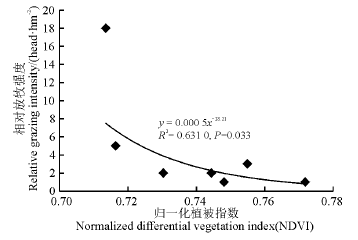 | 图6 调查点植被指数和相对放牧强度的幂回归模型(剔除特殊点之后)Fig. 6 Power regression model for vegetation index and relative grazing intensity (after exclusion of special points) |
在玛曲高寒草甸基于MODIS-NDVI的线性模型能更好地模拟高寒草甸地上生物量, 张艳楠等[18]发现当生物量低于370 g· m-2时, 建立的估产模型都是一元线性的, 当生物量在370~720 g· m-2时, 一元线性模型和指数模型的模拟效果都很好。本研究中生物量最大值为544.12 g· m-2, 比较发现线性模型模拟较好, 这与前人研究结果一致。罗媛等[19]在基于红光和近红外反射光谱特征参数反演草地地上生物量的研究中发现, 红光区域光谱特征参数与草地地上生物量之间存在显著的相关关系, 试验表明, 地上生物量的增加能够显著降低“ 红谷” 反射率, 显著升高近红外850 nm处反射率, 因此NDVI值随着生物量的增加而增大, 这就可以解释本研究中NDVI随着地上生物量的增加而增长的趋势。比较分析不同月份的地上生物量和NDVI值, 发现9月时生物量有所下降, 而且NDVI值也较6月、7月和8月有所下降。一般情况下, 牧草会在5月开始返青, 地上生物量从生长季开始逐渐增加, 到8月达到最大值, 而后进入非生长季, 牧草开始枯黄, 生物量的补偿生长滞后于放牧的影响, 放牧会影响其群落组成且降低禾本科牧草的生物量[20], 故到9月时地上生物量有所下降。而且, 因为红光波段是绿色植物进行光合作用吸收的主要波段, 9月地上生物量枯黄会导致光合作用减弱, 对红光波段的反射增强, 所以NDVI值较于6月、7月和8月会变小。此外, 模型的相关系数比较低, 可能是受研究区土壤、气候等条件的影响, 另外, 选取的MODIS产品数据空间分辨率有限, 也会影响估产模型的精度。
仁青吉等[21]研究结果表明, 随着放牧强度的增加, 青藏高原东部玛曲高寒嵩草草甸群落的高度、盖度和地上生物量都呈现显著降低的趋势。张伟华等[22]研究发现, 随着牧压强度的增加, 锡林河北岸一级台地退化草地的植物群落高度降低, 地表盖度下降, 地上生物量较大幅度减少, 特别是优质牧草生物量迅速减少。这与本研究结果一致。李建龙等[23]在新疆石河子紫泥泉种羊场进行的不同强度的分区轮牧试验也表明, 不同轮牧强度处理对草地地上部产量有显著影响。范国艳[24]研究发现, 随着放牧强度的增加, 贝加尔针茅典型草原区的贝加尔针茅(Stipa baicalensis)草原群落植物物种数、植被盖度、地上部现存量均显著降低, 植物群落种类组成结构发生显著变化。Joneydi等[25]研究发现, 虽然重度放牧降低了干旱灌丛带地上生物量, 但中度放牧对该区植物生物量没有影响。在本研究中, 中度放牧对该地区地上生物量是有一定影响的, 该差异可能与研究区域的植被类型、土壤和物候条件等因素有关。
不同水分条件下的不同植被群落类型对牧草生长速率有一定的影响。模型中提到的两个特殊点分别是牧户2和牧户3在重牧梯度下的NDVI值, 一般情况下, NDVI值随着放牧强度的增大而变小, 但此两点处NDVI值并没有变小, 究其原因发现, 牧户2和牧户3的牧场有河流经过(图2), 在沿河地区, 植被层保持着较高的含水量, 植物种类成分属于沼泽草甸, 而且植物生产力比周边草原地区的要高。因此放牧强度虽然较大, 其生物量也较高, 故NDVI值也较高。Kawamura等[26]也发现, 植物生物量随着放牧强度的增加而下降, 除了锡林沿河地区, 本研究的情况与该研究结果一致。作为家畜喜食, 但又不耐放牧践踏的植物, 如鹅绒委陵菜(Potentilla anserina)、羊茅(Festuca ovina)、垂穗披碱草(Elymus nutans)、草地早熟禾(Poa pratensis)等受到放牧干扰影响较大, 不能充分生长和发育, 其盖度和丰富度有所下降[27], 即植物的生成速率小于家畜的采食速率, 这可能是NDVI值随着放牧强度的增强呈下降趋势的原因之一。另外, 从整个群落角度分析, 放牧显著降低了地上生物量, 主要原因在于叶片光合面积的减少, 有机物积累效率的降低[28]。曹广民等[29]选取处于不同退化阶段的具有典型代表意义的草甸为研究对象, 通过对其土壤氮素矿化补给能力、牧草对氮素的需求量的研究, 发现随着放牧强度的增加, 土壤-牧草营养供求失调, 土壤养分被地下根系过多吸收被生物固定, 成为无效养分[30], 这也是生物量降低的主要原因之一, 也可能导致研究区域NDVI值的下降。分析相对放牧强度和NDVI之间的关系, 可以发现一个相对放牧强度的临界值, 在该值下, NDVI对相对放牧强度的响应不再灵敏, 这可以作为一个衡量放牧程度强弱的警戒值, 对于确定合理放牧强度具有重要的参考价值, 而且生物量随放牧强度的变化也有一个放牧强度阈值存在。草地地上生物量的直接收获估测方法需要花费相当大的时间和劳力, 同时要求足够多的样本数目, 以保证样本符合统计学的要求(样本数量少则估测精度有限)[31]。此外, 直接收获法还会破坏调查地点的植被, 具有破坏性, 对于一些生态脆弱区域的植被恢复有一定影响, 难以大面积开展, 再加上家畜的活动是空间动态变化的过程, 如果利用放牧强度和地上生物量的相关关系来寻找放牧强度阈值, 不仅工作量大, 而且有空间限制, 对研究结果有很大的影响。但是, 与基于样方的生物量监测的传统方法相比, 遥感估测不仅有更高的时效性, 而且具有传统方法无法比拟的优势, 如可以监测高寒无人区草地的变化动态等[32]。所以, 利用遥感植被指数来获取家畜的放牧强度阈值具有可行性。
本研究利用归一化植被指数(NDVI)和地面实测数据进行相关性分析, 结果发现玛曲高寒草甸不同月份的地上生物量与对应的植被指数有较好的相关性, 据此建立了6-9月份调查点植被指数和地上生物量的线性回归模型。发现6-9月的模拟呈现相同趋势, 即随着NDVI值的增长, 地上生物量呈现增长趋势。实测地上生物量和相对放牧强度有很好的拟合关系(R2=0.965 1), 随着放牧强度的增加地上生物量呈下降趋势。另外, 相对放牧强度和植被指数间相关性较好, 并建立了相对放牧强度估测模型(y=0.000 5x-28.21, R2=0.631 0), 结果表明随着NDVI值的变小, 相对放牧强度逐渐增强, 当相对放牧强度增强到一定程度时, NDVI对其响应不灵敏。量化这一程度上的放牧强度(即阈值), 是今后研究中需要做的工作。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|