


{kind=link}
{kind=link}
{kind=link}
三江源区高寒草原土壤湿度变化特征及与气候因子的关系
[郭连云 ]
]
]
|
|


利用青海省三江源区兴海县牧业气象站1999-2016年高寒草原土壤湿度、牧草生育期资料,分析了高寒草原土壤湿度的年、季变化特征以及牧草生长季不同生育期土壤湿度的变化特征及与气候因子的关系。结果表明:高寒草原0-10、10-20、20-30、30-40和40-50 cm土壤湿度均随年际延长呈增加趋势,春季除40-50 cm土层外,其他各层土壤湿度均随年际延长呈显著增加趋势( P<0.05)。高寒草原牧草生长季的土壤相对湿度随年际延长呈显著上升趋势( P<0.05),且与生长季降水量之间呈极显著正相关关系( P<0.01)。随着气候变化,牧草抽穗、开花、成熟和枯黄期的土壤相对湿度随年际延长呈显著增加趋势( P<0.05)。牧草抽穗期、枯黄和全生育期的土壤湿度与气温呈显著负相关关系( P<0.05)。高寒草原牧草生长季土壤湿度的增加有利于草地植被生长。
The data supporting this study were collected during 1999-2016 from the pastureland weather station of Xinghai County, Qinghai Province, at the Three River Source. They included soil moisture content and herbage growth period. We analyzed the changes of alpine meadow soil moisture characteristics, the soil moisture change characteristics at different stages, and their relationship with the herbage growth period. The results showed that the soil moisture content in each layer increased from 0-50 cm in the alpine grassland, the soil moisture was significantly increased trend of each layer in spring 0 to 40 cm, the soil moisture increase rate decrease with the increase of soil depth. Soil moisture content during growing seasons increased significantly from 1999-2016, and it correlated significantly with precipitations during the growing seasons. At the flowering stages, soil moisture content was significantly positively correlated with atmospheric temperature. Moreover, at the heading stages and throughout the whole growing season, they were significantly negatively correlated with atmospheric temperatures. The increase of soil moisture content in alpine meadow growth season is beneficial for grassland vegetation growth.
草地作为陆地的重要生态系统, 是生态环境的基础, 也是畜牧业发展的重要基础和牧民赖以生存的基本生产资料[1, 2, 3]。土壤水分是影响青藏高原高寒草地生态过程、生态承载能力以及退化草地恢复和重建的关键因素之一[4], 同时, 土壤水分对植物的生长期发育和产量的形成有直接作用[5]。对青藏高原多年冻土区高寒草地地上生物量影响最大的因子是土壤含水量[6]。土壤水分是主要生态因子之一, 是对气候因子变化最敏感的响应指标, 对于某一特定的区域而言, 土壤结构、地形、植被等自然因素在一定的时期、季节内是相对稳定的, 而气候因子的变化彼此间并非始终一致, 因而气象因子变化对土壤湿度波动起着决定性作用[7, 8, 9]。
众多研究[10, 11, 12, 13, 14, 15]表明, 干旱半干旱区自然降水是该区域土壤水分的主要补充源, 自然降水的变化直接影响土壤水分的变化, 从而影响植物水分利用策略, 进而影响整个生态系统的结构和功能。地下生物量与土壤含水量有显著的正相关关系, 在同样的温度条件下, 土壤含水量是地下生物量的重要影响因子[16]; 土层水分含量的好坏直接影响着植物的生长, 而且间接影响植物分布并在一定程度上影响小气候的变化[17, 18]。在旱作区土壤水分是作物所需水分的直接来源, 是作物生长发育和产量形成的关键因素[19, 20, 21]。以往的研究大多集中在通过短期观测数据来揭示土壤水分的动态变化, 本研究则侧重在利用长期观测的气象数据和土壤含水量数据来探讨高寒草原牧草生长季及各生育期间土壤湿度变化特征, 同时研究高寒草原气候因子与土壤湿度之间的关系, 以期揭示高寒草原土壤湿度变化规律, 从而实现高寒草原土壤湿度的充分开发利用, 为合理利用高寒草原土壤水资源及牧草的生产和决策提供科学依据。
研究区位于黄河上游“ 三江源” 地区的兴海县, 属于环湖牧区和青南牧区过渡的区域, 99° 01'-100° 20' E, 34° 48'-36° 14' N。全县总面积121.86万hm2, 平均海拔4 300 m, 全县地势西南高, 东北低, 境内地貌类型多样。草原面积101.03万hm2, 占全县总面积的83%, 可利用草地面积93.6万hm2, 占草地面积的92.7%。气候具有显著的高原大陆性气候特征, 年平均气温1.4 ℃, 气温极端最高30.2 ℃, 极端最低-31.5 ℃, 年降水量353.2 mm, 年日照时数4 431.8 h, 昼夜温差大, 年平均大风日数48.9 d, 年平均沙尘暴日数11.1 d, 无霜期44 d。草地类型为高寒草原天然草地, 草本植物以西北针茅(Stipa sareptana var. krylovii), 伴生冷地早熟禾(Poa crymophila)、斜茎黄芪(Astragalus adsurgens)、矮嵩草(Kobresia humilis)、猪毛蒿(Artemisia scoparia)等, 植被均匀, 覆盖率在60%以上。
1.2.1 样地设置与资料来源 样地位于兴海县子科滩镇高寒针茅草原, 地理位置为35° 35' N, 99° 59' E, 海拔3 300 m, 设立于1999年。样地面积为50 m× 50 m, 每年4月至9月开始观测牧草发育并围栏封育禁牧, 10月至次年3月期间放牧。依据该草原优势牧草西北针茅的生长变化, 按照中国气象局牧业气象观测规范, 观测牧草的返青、抽穗、开花、成熟、枯黄等5个生育期时间, 在围栏草地牧草生长发育期取4个重复样方, 每个样方面积1 m× 1 m, 4-8月份为牧草生长季节。选用兴海县牧业气象观测站采集的1999-2016年18年土壤湿度资料, 在春季土壤表层0-10 cm完全融化至土壤冻结深度≤ 10 cm期间的每旬逢8日测定土壤湿度, 采用土钻法分层取样, 分别为0-10、10-20、20-30、30-40和40-50 cm, 共5个层次, 各层均取4个重复, 每层重复取样4次, 然后用烘干法(105 ℃)烘干称重, 取其平均值为该层的土壤水分含量。测定方法是在天然牧草固定观测地段, 从4月8日开始测定水分, 测定时间间隔为10 d, 最后一次为10月28日, 共测定7个月, 采集21次。从距样地2.5 km处的兴海国家基准气候站获取同期的气温、降水、日照时数等观测资料。土壤相对湿度为土壤重量含水率占田间持水量的百分比。
土壤相对湿度=[土壤重量含水率(%)/田间持水率(%)]× 100%。
1.2.2 计算分析方法 采用线性气候倾向估计法[22]研究1999-2016年18年牧草生长季降水量及各生育期土壤相对湿度变化趋势。线性气候倾向估计法属于时间序列分析范畴, 该方法是对资料的时间序列y(x), x=1, 2, 3, ……, n, 以线性函数y(x)=a+bx来拟合, 其中, a为常数, b为回归系数, 用最小二乘法估计回归系数b和常数a。其中b为气候倾向率, 单位为每年某要素单位。回归系数b的正负表示该资料序列随时间变化的方向, 正值表示气象要素随时间变化有增多的趋势, 负值则表示有减少的趋势。采用气候要素与年份的相关系数来确定变化是否显著。
利用相关分析法[23]计算牧草各生育和同期影响牧草生育期的土壤相对湿度及温度、降水和日照时数的相关系数, 根据相关系数的大小, 判断土壤相对湿度、温度、降水和日照时数等与牧草各生育期是否有明显相关关系。采用Excel 2003和DPS 7.05进行数据整理、制图和统计分析。
2.1.1 土壤湿度年变化特征 0-10、10-20、20-30、30-40和40-50 cm土层土壤湿度均随年际呈增加趋势, 且在2009年达到最高值(图1)。最低值除10-20、40-50 cm分别出现在2013、1999年外, 其余各层均出现在2000年。0-10、10-20、20-30、30-40和40-50 cm各层土壤湿度最高值与最低值之差分别为48.7%、34.6%、30.7%、26.8%、14.9%, 可见随着土壤深度的增加其差值逐渐减少。0-50 cm土壤湿度最高值与最低值的差值为31.1%, 变化幅度较大。18年间0-50 cm土壤湿度总体呈现逐渐上升的变化趋势, 将其变化趋势可以划分为两个阶段:第1阶段为1999-2009年, 该阶段土壤湿度年均值在62.3%~93.4%之间波动, 属于土壤湿度的较高阶段; 第2阶段为2000-2016年, 该阶段土壤湿度年均值在67.6%~83.8%之间波动, 属于土壤湿度的较低阶段。各层土壤湿度的各阶段变化趋势和0-50 cm土壤湿度的基本一致。
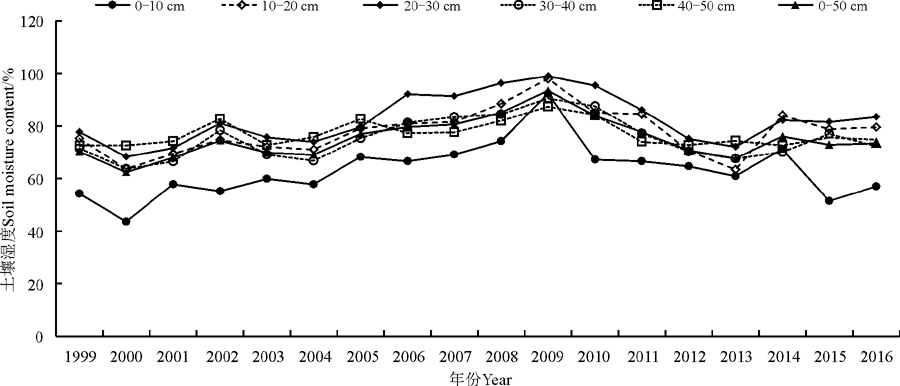 | 图1 土壤湿度年变化趋势Fig. 1 The annual trend of soil moisture content |
高寒草原0-10、10-20、20-30、30-40和40-50 cm土层土壤湿度的气候倾向率分别为0.60%· 年-1、0.55%· 年-1、0.54%· 年-1、0.33%· 年-1和0.00%· 年-1, 各土层土壤湿度与年份的相关系数分别为0.303(P> 0.10)、0.335(P> 0.10)、0.310(P> 0.10)、0.222(P> 0.10)和0.003(P> 0.10), 随着土壤深度的增加, 其气候倾向率逐渐减小, 到40-50 cm时气候倾向率无增无减, 变化平稳(表1)。而0-50 cm整层土壤湿度则以每年0.40%的气候倾向率增加, 土壤湿度与年份的相关系数为0.287(P> 0.10), 但增加趋势小于0-10、10-20和20-30 cm土壤湿度。虽然各土层土壤湿度随年际延长均呈增加趋势, 但增加趋势均未通过显著性检验, 说明各层土壤湿度随年际延长增加的趋势不明显。
| 表1 春、夏、秋季和年土壤湿度的年气候倾向率及其与年份间的相关系数(n=18) Table 1 The limate tendency rate and the correlation coefficient of the soil moisture content in spring, summer, autumn, and throughout the year |
2.1.2 土壤湿度季节变化特征 由于测定的是3月中旬至11月上旬解冻到封冻时段的土壤湿度, 所以本研究只分析春、夏、秋季土壤湿度的变化。春季高寒草原0-10、10-20、20-30、30-40 cm各层土壤湿度随年际延长均呈显著增加趋势, 其气候倾向率在0.85%~1.21%· 年-1, 而40-50 cm土壤湿度的气候倾向率为0.46%· 年-1(P> 0.10), 增加不显著(表1)。春季土壤湿度与年变化一样随着土壤深度的增加其气候倾向率总体呈逐渐减小趋势。而春季0-50 cm土壤湿度则以每年0.88%速率显著增加, 相关系数为0.557(P< 0.05)。
夏、秋两季中, 40-50 cm土壤湿度均呈减少趋势, 但不显著(P> 0.05), 其中秋季的减少速度大于夏季。0-10、10-20、20-30、30-40 cm各土层土壤湿度在夏、秋季均呈不显著的增加趋势, 其中各土层夏季的增加幅度表现为0-10> 20-30> 10-20> 30-40 cm; 秋季的增加幅度表现为20-30> 10-20> 0-10> 30-40 cm。
兴海高寒草原牧草全生育期0-50 cm土壤相对湿度和降水量随年际延长的线性变化趋势如图2所示, 可见兴海高寒草原牧草生长季土壤相对湿度以每年0.51%的线性速率增加, 土壤相对湿度与年份的相关系数为0.380(P> 0.10)。牧草生长季土壤相对湿度多年平均值为75%, 最小值为62%, 最高值为91%, 18年中有9年高于多年平均值, 2004-2009年土壤相对湿度增加最快, 2010年开始又急剧下降, 2013年以后土壤相对湿度又呈现出增加趋势。牧草生长季降水量则以每年3.5 mm(P> 0.10)的线性趋势呈不显著的增加趋势。牧草生长季降水量波动变化较大, 多年平均值是326.5 mm, 2000年为最少年(205.8 mm), 2005年为最多年(441.2 mm), 最多年与最少年之间相差2.14倍, 年际间差异显著(P< 0.05)。牧草生长季土壤相对湿度和降水量的年变化均呈增加趋势, 二者之间极显著相关(P< 0.01)。可见, 自然降水是近18年来影响兴海县高寒草原牧草生长季土壤干湿程度的主要因素。
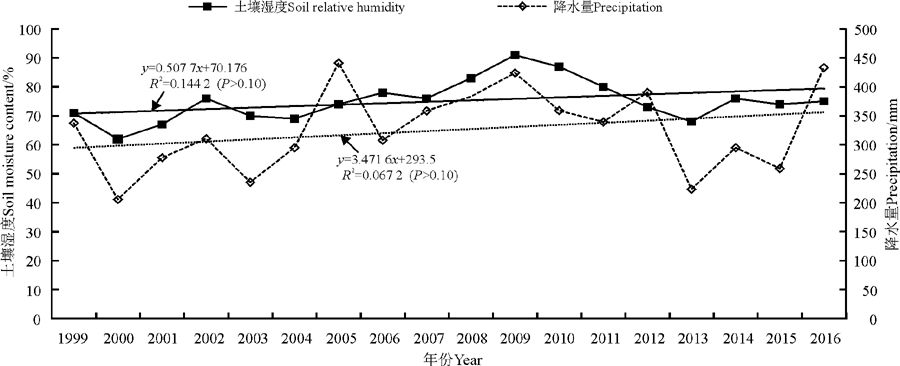 | 图2 牧草全生育期土壤相对湿度和降水量的变化Fig. 2 Change in of soil moisture content and soil relative humidity during the whole growing season of the pasture |
高寒草原牧草返青期土壤相对湿度以每年0.77%的线性趋势增加, 其土壤相对湿度与年份的相关系数为0.315(P> 0.10)。成熟和枯黄期的土壤相对湿度分别以每年1.06%和1.70%的线性速率增加, 土壤相对湿度和年份的相关系数分别为0.567(P< 0.05)、0.482(P< 0.05), 可见其随年际的增加趋势是显著的。而抽穗和开花期的土壤相对湿度分别以每年1.26 %和1.42 %的线性趋势增加, 其土壤相对湿度与年份的相关系数分别为0.609(P< 0.01)和0.651(P< 0.01), 可见抽穗和开花期土壤相对湿度随年际延长均呈极显著增加趋势(图3)。牧草各发育期中, 以枯黄期土壤相对湿度的线性增加速率最大, 返青期土壤相对湿度的线性增加速率最小。各发育期土壤相对湿度的大小依次为枯黄期> 开花期> 抽穗期> 成熟期> 返青期。
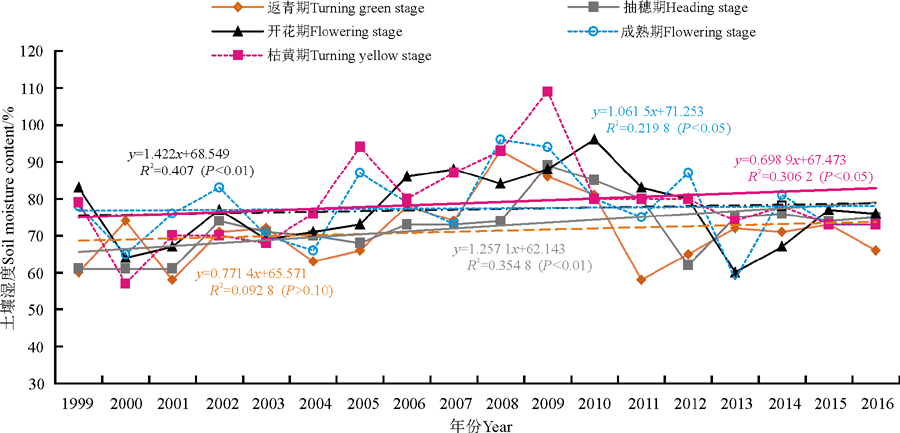 | 图3 牧草不同生育期土壤相对湿度变化Fig. 3 Change in soil moisture content during different growth periods |
兴海高寒草原牧草返青至枯黄期各生育期的平均土壤相对湿度在71%~79%, 变化幅度依次是:返青期在58%~93%, 抽穗期在61%~89%, 开花期在60%~96%, 成熟期在58%~96%, 枯黄期在57%~109%。从变化幅度可见, 牧草枯黄期间的土壤相对湿度最大, 开花和成熟期其次, 返青和抽穗期间的土壤相对湿度最小。
对高寒草原1999-2016年牧草生长季土壤湿度与降水、气温、日照等气候因子进行相关性分析(表2), 结果表明, 气温与牧草抽穗期(P< 0.10)、枯黄期(P< 0.10)和全生育期(P< 0.05)土壤湿度之间呈负相关关系。降水量与牧草抽穗期、开花期土壤相对湿度均呈显著正相关关系(P< 0.05); 成熟期和全生育期土壤相对湿度与降水量均为极显著正相关关系(P< 0.01)。枯黄期土壤相对湿度与日照时数呈负相关关系(P< 0.10)。
| 表2 土壤湿度与气候因子相关系数(n=18) Table 2 Correlation of soil moisture content with climate factors |
进入春季, 随着气温回升, 当气温达到牧草萌芽返青的界限温度时, 牧草开始萌芽返青, 土壤水分自上而下融化, 因此该阶段土壤湿度与气温之间表现为正相关关系。该阶段尽管气温回升快, 但自然降水很少(仅为年降水量的3.5%), 因此土壤湿度不但不增加, 反而增大了土壤水分的蒸发消耗, 造成土壤湿度降低。这是因为土壤水分的可利用性是牧草返青的决定因素, 但作为区域土壤水分唯一来源的降水主要集中在植物生长季, 非生长季几乎无降水, 返青期的早晚受控于生长季降水[24]。到6月上中旬时, 进入牧草抽穗期, 由于气温回升快, 植物生长速度加快, 植被盖度扩大, 导致耗水量增加, 此阶段该地易出现少雨时段, 造成不能有效补偿土壤水分, 土壤湿度不升反降, 因此表现出该时期气温与土壤湿度之间呈显著负相关关系, 而降水量与土壤湿度之间显著正相关。至7月中旬时, 牧草为开花期, 由于此阶段为当地主汛期, 降水过程及降水量明显增多, 有效补充了土壤水分, 土壤湿度较高, 水热条件满足牧草的生长发育及光合作用, 表现出气温和降水因子与土壤湿度之间显著正相关。土壤水分的有效性是草原植被的重要限制因子, 尤其区域植被多以浅根植物为主, 更加剧了种间水分的竞争。在牧草生长旺盛期, 也难以满足植物群落的需求, 加之区域雨热同期, 反应出土壤湿度和温度、降水显著正相关[25]。8月中旬为牧草的成熟期, 此阶段温度高、降水量多, 虽然相应的植物蒸腾和土壤蒸发量也较大, 但由于土壤浅层水分容易得到有效补给, 土壤水分较高, 表现出此阶段土壤湿度与降水量具有显著的正相关关系。9月上中旬时牧草进入枯黄期, 大部分植物由于完成了整个生育周期而基本停止生长, 此时土壤水分的消耗主要以土壤蒸发为主[26], 无论是土壤湿度与降水正相关关系, 还是土壤湿度与温度、日照时数之间负相关关系均不显著, 由于受到前期降水的缓慢累积, 降水对土壤湿度影响较小。气温与牧草全生育期土壤湿度显著负相关, 表明牧草生长季温度升高会加大土壤蒸发、作物蒸腾, 从而降低土壤含水量。由此可见, 降水季节分配是土壤水分季节波动的主要影响因素[27, 28]。由于近年来三江源大部分地区生长季降水量增加, 整体气候适宜程度呈上升趋势, 有利于草地植被生长[29]。
1)高寒草原0-50 cm各层土壤湿度随年际延长均呈增加趋势, 牧草生长季降水量波动变化是导致土壤湿度的升降变化的主要原因。2)高寒草原牧草生长季的土壤湿度随年际延长呈显著增加趋势, 且与降水量之间呈极显著正相关关系。3)气温与牧草抽穗期、枯黄期和全生育期的土壤湿度呈显著负相关关系。降水量与牧草抽穗期、开花期、成熟期和全生育期的土壤湿度均呈显著正相关关系。
(责任编辑 苟燕妮)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|