


{kind=link}
{kind=link}
{kind=link}
短时高温处理对意大利蝗卵子发生期HSP70蛋白表达的影响
[向敏, 扈鸿霞, 于非, 季荣, 王晗 ]
]
]
|
|


第一作者:向敏(1992-),女,新疆库尔勒人,在读硕士生,研究方向为昆虫生殖生态学。E-mail:1441109005@qq.com
本研究采用免疫组织化学分析方法,测定了HSP70蛋白在意大利蝗卵子发生期的表达定位及33~42 ℃高温处理对其相对表达量的影响。结果表明,1)在卵黄发生前期,卵母细胞和滤泡细胞均表达HSP70蛋白,而在卵黄发生期和卵黄发生后期,HSP70蛋白阳性表达位于滤泡细胞;2)在33~42 ℃温度范围内,随着温度升高,意大利蝗卵子发生期HSP70蛋白相对表达量呈现先上升后下降的变化趋势;其中36 ℃处理组HSP70蛋白相对表达量最高,显著高于27 ℃对照组( P<0.05);42 ℃处理组HSP70蛋白相对表达量最低,但与27 ℃对照组差异不显著( P>0.05)。短时高温处理对意大利蝗卵子发生期HSP70蛋白相对表达量有显著影响,推测HSP70蛋白的表达在意大利蝗抵抗短时高温胁迫过程中具有重要作用。
The expression of heat shock protein 70 (HSP70) in the oogenesis of Calliptamus italicus was detected by immunohistochemistry after the exposure of adult insects to temperatures of 33, 36, 39, and 42 ℃ for 4 h. The results showed that HSP70 was expressed in the oocytes and follicular cells during the previtellogenic stage, but was only expressed in the follicular cells only during the vitellogenic and postvitellogenic stages. The relative expression of HSP70 showed an initial increase, but then decreased with an increase in temperature in the range from 33~42 ℃. The highest expression, which was significantly higher than those in the control (27 ℃), was observed when adult insects were exposed to 36 ℃ ( P<0.05). The lowest expression was observed when adults were exposed to 42 ℃, but there was no significant difference between treatment at 42 and 27 ℃ ( P>0.05). The relative expression of HSP70 was significantly different at different temperatures, which may be critical for the resistance and adaptation of C. italicus to brief exposure to high temperature.
近年来, 全球气候变暖的趋势加剧, 极端高温发生的频率增加[1, 2]。昆虫是变温动物, 对环境温度高度敏感, 极端高温的出现对其生长发育、生理代谢及生殖等具有重要影响[2, 3]。但是, 田间高温可能仅在某一天发生且仅持续几个小时, 而且昆虫在长期进化过程中, 形成了较强的适应能力以应对短期环境变化[4], 偶尔的高温不会导致其死亡。然而, 大量研究表明, 昆虫的生殖系统对高温极其敏感, 因此极端高温的出现, 可能对昆虫的生殖系统造成影响[5, 6, 7]。
热激蛋白(heat shock proteins, Hsps)的表达和调控是有机体应对高温胁迫的物质基础之一[8, 9]。HSP70是热激蛋白家族的主要成员, 在提高机体耐热性方面具有重要作用。昆虫在受到高温胁迫后, 其耐热能力与HSP70的表达量正相关, 且在一定温度范围内, HSP70表达量随温度升高而增加[10, 11, 12, 13]。此外, 热激蛋白家族与有机体生殖系统发育密切相关。侯赛尼[14]对热应激后家蚕卵巢和精巢进行检测发现, HSP70表达上调。对哺乳动物研究表明, HSP70不仅参与精子发生和成熟过程, 而且在卵子发生和卵母细胞存活方面具有重要作用[15]。
配子发生是决定昆虫生殖力的基础, 是昆虫种群繁衍和延续的重要保障。气候变暖背景下, 害虫始终保持较高的产生配子的能力, 是其持续发生并暴发成灾的重要原因[16, 17, 18, 19]。其中, 卵子发生在动物的个体发育过程中起着至关重要的作用, 一般分为3个时期:卵黄发生前期、卵黄发生期及卵黄发生后期。卵黄发生是昆虫生殖调控的核心[20, 21], 包括脂肪体合成卵黄原蛋白(vitellogenin, Vg)、卵巢摄取并沉积卵黄蛋白(vitellin, Vn)两个主要过程。卵母细胞中卵黄蛋白的迅速积累, 可促进卵母细胞和卵巢的发育成熟, 亦可为胚胎发育提供营养物质, 有利于提高孵化率及幼虫生长发育, 对昆虫种群数量有显著影响[22]。
意大利蝗(Calliptamus italicus)是新疆荒漠半荒漠草原的优势危害种类, 每年给新疆的畜牧业经济、生态和社会造成严重的损失[23, 24]。近年来, 意大利蝗适生区气温升高明显[25], 繁殖期(5月-8月)日极端高温经常会超过33 ℃, 甚至出现42 ℃极端高温, 且极端高温出现天数也在逐年增加[26]。田间短时高温是否对意大利蝗生长繁殖造成影响?为此, 本研究采用免疫组织化学分析方法, 研究短时高温处理对意大利蝗卵子发生期HSP70表达的影响, 在细胞水平上直观分析HSP70蛋白在卵子发生中的准确定位及表达动态, 以期为进一步探讨温度胁迫下意大利蝗卵子发生中HSP70蛋白在细胞水平的作用机理及分子调控机制奠定基础, 同时阐明意大利蝗应对高温胁迫时HSP70在生殖系统中的调控作用, 为在细胞水平上揭示蝗虫生态适应的耐热机理提供科学依据。
于新疆哈密白石头乡(93° 80' E, 43° 31' N)采集意大利蝗末龄蝗蝻, 置于室外(100 cm× 100 cm × 100 cm养虫笼)饲养, 待成虫羽化后, 取同天羽化的成虫, 置于光照培养箱内继续饲养[30 cm× 30 cm × 30 cm养虫笼, (27± 1) ℃, 相对湿度(45%± 5%), 光照周期L∶ D = 14∶ 10], 以新鲜玉米叶、小麦叶饲喂, 每天更换玉米叶与小麦叶。
根据前期试验可知, 意大利蝗羽化后10~15 d卵巢管内存在各个时期的卵母细胞, 故选取羽化后10~15 d蝗虫进行温度处理试验。 对照组:将羽化后成虫置于(27± 1) ℃、RH(45%± 5%)、光照周期L∶ D = 14 ∶ 10条件下常规饲养。
试验组:设置33、36、39、42 ℃ 4个温度处理组, 每个温度梯度处理4 h, 处理后将其放回(27± 1) ℃、相对温度(45%± 5%)条件下恢复1 h。每个温度处理15头雌虫, 3个重复。
在0.65%生理盐水中进行解剖取出卵巢, 放入Bouin’ s液中固定24 h, 常规石蜡包埋与切片, 切片厚度为6 μ m。 将切片常规脱蜡入水, 用0.01 mol· L-1枸橼酸盐缓冲液(pH 6.0)于96 ℃修复抗原20 min, 使其自然冷却; 加内源性过氧化酶阻断剂, 室温下孵育10 min, 以阻断内源性过氧化物酶活性; 经5%牛血清蛋白BSA室温封闭20 min; 滴加HSP70一抗(1∶ 100)(设滴加PBS组为阴性对照), 4 ℃过夜; 滴加生物素标记的第二抗体(山羊抗兔IgG), 37 ℃处理20 min; 滴加链霉亲和素-生物素(SABC)复合物37 ℃处理20 min, 增敏二氨基联苯胺(DAB)室温显色, 苏木精复染, 中性树胶封片[27]。显微镜下观察和拍照(Motic数码显微镜BA310)。
免疫组织化学染色后棕黄色区域为阳性区域。不同温度处理下随机取10张片子, 每张切片随机选取3个视野, 使用Image-pro plus 6.0专业图像分析软件分析HSP70相对表达量。
采用SPSS 18.0统计软件对数据进行分析, 用平均值和标准误表示测定结果, 不同温度处理组间HSP70相对表达量进行单因素方差分析, 并采用Duncan法对各测定数据进行多重比较。采用Excel 2013制图。
意大利蝗卵巢结构为无滋式, 只有卵母细胞和滤泡细胞, 无滋养细胞。在生长区内含有由大到小的卵母细胞, 每个正在发育的卵母细胞被一层滤泡细胞包围。参照赵卓[28]对10种蝗虫卵子发生的划分, 根据意大利蝗卵母细胞和滤泡细胞形态及变化特征, 将意大利蝗卵子发生分为3个时期, 分别为卵黄发生前期、卵黄发生期和卵黄发生后期。免疫组织化学分析显示, 在卵黄发生前期, 卵母细胞和滤泡细胞均呈现HSP70蛋白阳性表达, 而在卵黄发生期和卵黄发生后期, HSP70蛋白阳性表达仅位于滤泡细胞(图1)。
 | 图1 意大利蝗卵子发生期HSP70的特异性表达Fig. 1 Specific expression of HSP70 in the oogenesis of Calliptamus italicus |
免疫组织化学结果表明, 不同温度下意大利蝗卵子发生期均有HSP70蛋白表达(图2)。对免疫组织化学结果进行图像分析后表明, 短时高温处理对意大利蝗卵子发生期HSP70蛋白相对表达量有显著影响, 随着温度升高, HSP70蛋白相对表达量呈现先上升后下降的变化趋势(图3)。在卵子发生不同时期, 33、36、39 ℃处理组HSP70蛋白相对表达量均显著高于对照组(27 ℃)(P< 0.05), 其中36 ℃处理组HSP70蛋白相对表达量最高, 卵黄发生前期、卵黄发生期和卵黄发生后期分别为0.739、0.124和0.054。在卵子发生不同时期, 42 ℃处理组HSP70蛋白相对表达量低于对照组(27 ℃), 但差异不显著(P> 0.05)(图3)。
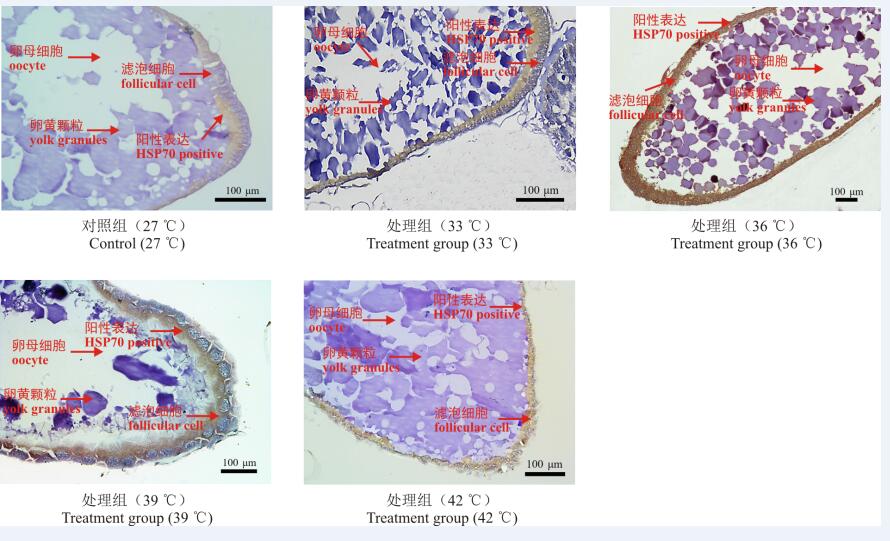 | 图2 不同温度下HSP70蛋白表达的免疫组织化学分析Fig. 2 Immunohistochemical assay of HSP70 expression at different temperatures |
 | 图3 意大利蝗卵子发生期HSP70相对表达量 注:不同小写字母表示不同温度处理间差异显著(P< 0.05)。Fig. 3 The relative expression level of HSP70 positive in oogenesis of Calliptamus italicus Note:Different lowercase letters indicate significant difference among different temperatures at the 0.05 level. |
意大利蝗卵子发生过程中, 随着卵母细胞和滤泡细胞形态、大小及功能的变化, HSP70蛋白的定位及相对表达量亦发生显著变化。在卵黄发生前期, 卵母细胞和滤泡细胞均处于旺盛生长期, 在高温胁迫下, 卵母细胞和滤泡细胞均大量合成HSP70以保证卵子发生和发育。在卵黄发生期, HSP70蛋白阳性表达仅位于滤泡细胞, 卵母细胞内未检测到HSP70蛋白的表达, 且滤泡细胞分泌合成的HSP70蛋白并不向卵母细胞中移动, 只停留并分布在滤泡细胞周围, 这可能是由于在卵黄发生期, 卵母细胞主要进行卵黄蛋白的积累以保证机体的繁殖力[28], 故仅由滤泡细胞合成HSP70蛋白抵抗高温胁迫。在卵黄发生后期, 由于滤泡细胞逐渐退化, HSP70蛋白相对表达量减少。
热激蛋白是普遍存在于原核和真核生物中的一类高度保守的蛋白, 其表达不仅有助于昆虫耐热性的获得, 且在昆虫的生殖、发育等方面起着一定的作用[29, 30, 31]。本研究表明, 对照组和处理组意大利蝗卵子发生期的滤泡细胞均可表达HSP70蛋白, 且在适宜生长温度(27 ℃)、旺盛生长温度(33、36 ℃)和热胁迫温度(39、42 ℃)下[32, 33, 34], HSP70蛋白相对表达数量呈现出先增后减的趋势, 表明意大利蝗体内能够大量合成热激蛋白以应对高温胁迫, 从而对机体起重要的保护作用[35]。20~30 ℃是意大利蝗生长发育适宜的温度条件[33], 故在此温度范围对意大利蝗并不构成胁迫, 细胞内表达的HSP70蛋白与热应激无关, 主要为生理条件下表达的组成型HSP70蛋白[36, 37]。意大利蝗成虫在29~35 ℃生长最为旺盛[32], 为促进卵子发生及卵巢发育, 机体可能大量合成组成型HSP70蛋白。同时, 高于30 ℃的温度条件对意大利蝗造成一定热胁迫, 机体可能合成大量诱导型HSP70蛋白以提高其耐热性。组成型HSP70蛋白和诱导型HSP70蛋白的累积, 可能是33和36 ℃处理组HSP70蛋白相对表达量显著增加的主要原因。当温度升高至39和42 ℃时, HSP70蛋白相对表达量显著下降(P< 0.05), 表明HSP70蛋白的热保护作用只能在一定温度范围内起作用[11, 27, 38]。但39、42 ℃处理组意大利蝗存活率仍为100%(数据未发表), 这可能是由于除HSP70蛋白外, 机体仍能通过合成热激蛋白家族其他蛋白以增强机体耐热性[39, 40, 41]。
本研究表明, 在一定温度范围内, 高温胁迫可引起意大利蝗卵子发生期HSP70蛋白表达增强, 以此提高其生殖系统的高温耐受性。HSP家族中HSP70对环境胁迫最为敏感[42, 43]。在温度胁迫条件下, 细胞内出现大量变性蛋白, 而HSP70蛋白作为一种非特异性保护蛋白, 可帮助变性蛋白恢复正确构象, 从而增强生物的耐热能力, 这可能是热激蛋白在胁迫细胞内的重要保护机制[44]。
(责任编辑 武艳培)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|