

{kind=link}
{kind=link}
盐旱胁迫对花花柴种子萌发与幼苗生理生化特性的影响
[石新建1, 2 , 王彦芹1, 3 , 李志军1, 3  ]
]
]
|
|


第一作者:石新建(1989-),男,山东滕州人,助理研究员,硕士,主要从事植物逆境生理与基因工程研究。E-mail:670648411@qq.com
为探明花花柴( Karelinia caspica)种子萌发和幼苗对盐旱胁迫的耐受性,采用不同浓度的PEG-6000溶液和NaCl溶液模拟盐旱胁迫,研究盐旱胁迫对花花柴种子萌发及幼苗保护酶活性、相对电导率和丙二醛(MDA)含量的影响。结果表明,1)花花柴种子最终萌发率随着NaCl和PEG胁迫程度的升高而降低,低浓度胁迫下的种子最终萌发率与对照无显著差异( P>0.05);当PEG质量分数大于20%、NaCl溶液浓度大于100 mmol·L-1时,各处理下的种子最终萌发率差异显著( P<0.05)。2)NaCl浓度为200~300 mmol·L-1和PEG质量分数为5%~15%胁迫下的花花柴幼苗叶片丙二醛(MDA)含量、过氧化物酶(POD)活性、超氧化物歧化酶(SOD)活性和过氧化氢酶(CAT)活性随着胁迫时间延长逐渐升高;当幼苗在400~500 mmol·L-1NaCl和25%的PEG胁迫下时,叶片MDA含量随着胁迫时间延长逐渐升高;POD、SOD和CAT活性随着胁迫时间延长先升高后降低。可见,花花柴对逆境环境具有较强的适应能力;25% 的PEG为其耐旱临界值,300 mol·L-1NaCl为其耐盐临界值。
In order to explore the physiological changes in Karelinia caspia during seed germination and seedling development under drought and salt stress, we used different concentrations of PEG-6000 or NaCl to simulate drought and salt stress, respectively. Germination rate, protective enzyme activity, relative conductivity and the Malondialdehyde (MDA)content during seed germination and in the seedling of K. caspia under stress (drought and salinity) were determined. The results show that: 1) With increasing amounts of PEG and NaCl, a gradual decrease in germination was observed, while there were no significant differences between the seeds treated with 15% PEG and 50 mmol·L-1 NaCl when compared to control. However, when PEG was above 20%, or when NaCl concentration was increased above 100 mmol·L-1, we observed significant differences between treated samples and control. 2) In the range of 5%~20% PEG or 200~400 mmol·L-1 NaCl, with a concentration gradient and increasing stress duration, a corresponding increase in MDA content, and elevated enzymatic activities for POD, SOD and CAT were observed. When plants were treated with 25% PEG and 500 mmol·L-1 NaCl, the relative conductivity and MDA content increased significantly with prolonged stress periods. Enzyme activities for POD, SOD and CAT also increased initially with prolonged stress before decreasing to normal levels. These preliminary results reveal that K. caspia Less has huge potential to adapt to adverse environmental conditions. Also, our study revealed that the drought threshold was equivalent to the PEG 25%, while the salinity threshold was the equivalent of 300 mmol·L-1 NaCl.
全球干旱和半干旱土地的面积占陆地总面积的30%以上, 20多个国家和地区均有分布。我国现有荒漠化土地总面积为261.16万 km2, 占国土总面积的27.20%, 其中重度、极重度荒漠化土地面积为93.68万 km2。我国北方地区的干旱、半干旱土地面积161.71万 km2, 占国土总面积的16.85%[1]。干旱、半干旱地区沙质土地的不合理利用, 往往会引起沙漠和戈壁面积的扩大以及土壤次生盐渍化, 严重影响陆地生态系统的结构和功能[2]。土壤中水分和盐分的含量不仅直接影响植物的种子萌发和生长, 而且影响植物体内丙二醛、抗氧化酶、叶绿素、糖和脯氨酸等有机物大分子的生理生化调控[3, 4]。
花花柴(Karelinia caspica)属菊科(Compositae)花花柴属多年生草本植物。在我国主要分布于新疆准噶尔盆地和塔里木盆地、青海柴达木盆地、甘肃西北部和北部、内蒙古西部, 多大片群生于干旱、半干旱地区河谷冲积平原及沙质草甸盐土上[5]。花花柴作为重要的防风固沙植物, 抗风蚀、耐沙埋、耐旱和耐盐碱, 具有很强的生态适应性, 对于改良干旱、半干旱退化草地和水土保持具有重要作用[6]。因此, 本研究对花花柴种子和幼苗进行持续干旱、盐胁迫试验, 对胁迫下的花花柴种子萌发与幼苗生理生化特性进行研究, 旨在评价花花柴耐逆特性, 为后续进一步从生理水平揭示其响应盐旱胁迫的调控机理奠定基础, 并为荒漠地区植被恢复与重建提供科学依据。
花花柴种子于阿拉尔市(40° 32' N, 81° 17' E)的荒漠土壤采得, 种子经处理后置于纸袋内, 4 ℃通风保存。
设置PEG质量分数为0、5%、10%、15%、20%、25%和30%模拟干旱, 在花花柴种子萌发最适温度和光照条件下(光照/黑暗20 ℃/25 ℃、16 h/8 h)进行种子萌发期渗透胁迫处理试验(试验期间每天换一次处理液)。每处理50粒种子, 3次重复。
设置NaCl浓度为0、50、100、150、200、250和300 mmol· L-1, 在花花柴种子萌发最适温度和光照条件下(光照/黑暗20 ℃/25 ℃、16 h/8 h)进行种子萌发期盐胁迫处理试验(试验期间每天换一次处理液)。每处理50粒种子, 3次重复。
种子萌发试验在人工智能气候箱中进行。种子萌发以胚根出现为标志, 每24 h检测一次种子萌发个数。种子萌发试验以连续3 d不萌发为萌发结束。
种子最终萌发率=处理15 d后萌发的种子数/种子总数× 100%。
将花花柴种子播种于土壤水分含量适宜的营养土中, 每个花盆播种15粒左右, 共播种150盆。待大部分种子萌发后, 每周灌水1~2次并浇透。室温培养温度为(23± 2) ℃, 光照16 h· d-1, 光照强度为600 μ mol· (m2· s)-1。待幼苗生长60 d时, 选取约高15 cm大小均匀的幼苗进行水培。
PEG胁迫:将水培花花柴幼苗分别移入质量分数为0、5%、10%、15%、20%、25%和30%的PEG-6000溶液中, 每处理5~7株幼苗, 重复3次。
NaCl胁迫:将水培花花柴幼苗分别移入0、200、300、400和500 mmol· L-1的NaCl溶液中。每处理5~7株幼苗, 重复3次。
各PEG和NaCl溶液浓度分别处理6、12、24和48 h后采集幼苗叶片, 分别测定相对电导率、丙二醛(MDA)含量、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)活性的生理生化指标, 每个指标3次重复。MDA含量采用硫代巴比妥酸法[7]测定; SOD活性采用氮蓝四唑(NBT)法[8]测定; CAT活性采用紫外吸收法[7]测定; POD活性采用愈创木酚法[7]测定。
试验数据展示为3次重复试验的平均值± 标准差。利用利用Microsoft Excel 2010软件进行数据分析和作图, 应用SPSS 17.0统计软件进行方差分析及LSD检验(P< 0.05)。
2.1.1 盐旱胁迫对花花柴种子最终萌发率的影响 NaCl胁迫处理时, 花花柴种子在对照下(0 mmol· L-1)的萌发率为91.33%, 随着NaCl浓度的升高种子最终萌发率逐渐降低。当浓度≥ 100 mmol· L-1时, 各浓度处理下的种子萌发率与对照差异显著(P< 0.05), 50 mmol· L-1处理下的种子最终萌发率小于对照, 但无显著差异(P> 0.05)(图1)。随着PEG质量分数的升高, 花花柴种子最终萌发率呈降低趋势。当PEG质量分数在20%~30%时, 各处理种子最终萌发率显著小于对照(P< 0.05), 且各处理间种子最终萌发率差异显著, 当PEG质量分数在5%~15%时, 各处理间种子最终萌发率与对照无显著差异。
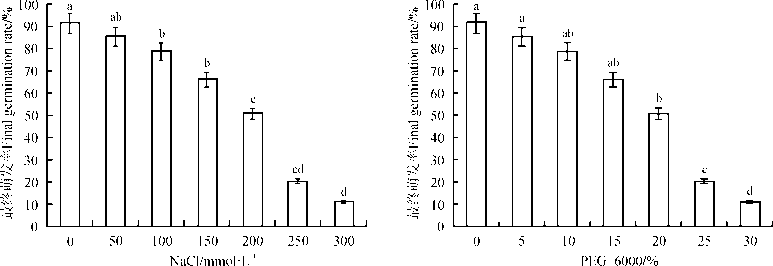 | 图1 盐、旱胁迫对花花柴种子最终萌发率的影响 注:不同小写字母表示处理间差异显著(P< 0.05)。Fig. 1 Effect of salt and drought stress on final germination rate of Karelinia caspia seed Note: Different lowercase letters indicate significant difference among different treatments at the 0.05 level. |
2.1.2 盐旱胁迫对花花柴种子萌发进程的影响 不同质量分数的PEG和不同浓度的NaCl溶液处理下的花花柴种子萌发集中在1~10 d, 胁迫加剧, 萌发进程越缓慢(图2)。低浓度胁迫下(PEG质量分数0~15%、NaCl溶液0~150 mmol· L-1)的种子萌发进程与对照类似, 高浓度胁迫下(PEG质量分数25%~30%、NaCl溶液200~300 mmol· L-1)的花花柴种子萌发表现出萌发时间晚、累计萌发率低的特点。
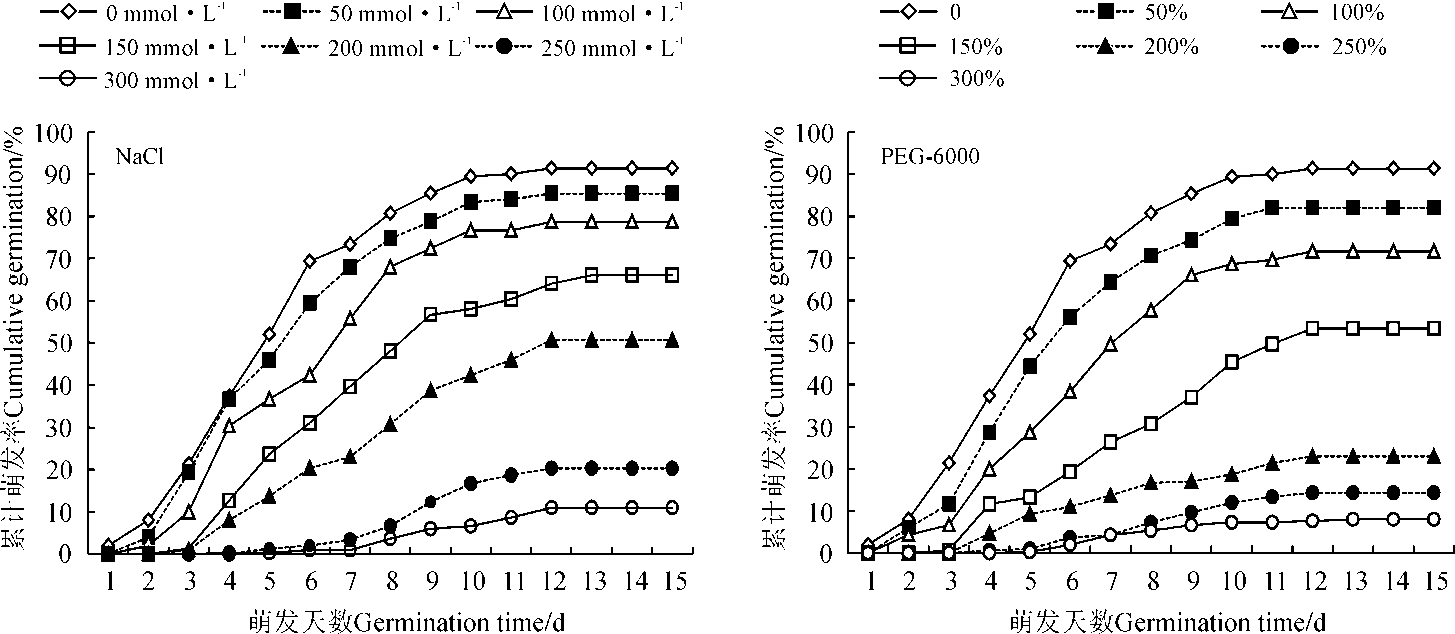 | 图2 盐旱胁迫对花花柴种子萌发进程的影响Fig. 2 Effect of salt and drought stress on germination process of Karelinia caspia seed |
2.2.1 盐旱胁迫对花花柴叶片丙二醛含量的影响 MDA含量是直接反映植物在逆境条件下细胞膜脂过氧化程度的重要指标之一。不同PEG和NaCl胁迫浓度和胁迫时间对花花柴幼苗叶片MDA含量影响不同(表1)。在低浓度胁迫下(NaCl溶液为200~300 mmol· L-1、PEG质量分数为5%~15%), 胁迫6~48 h下的幼苗MDA含量彼此无显著差异(P> 0.05), 且在胁迫12 h后MDA含量趋于稳定。当NaCl浓度≥ 400 mmol· L-1, PEG质量分数≥ 20%时, MDA含量随着胁迫浓度及胁迫时间的增加而升高, 除500 mmol· L-1胁迫外, 胁迫时间≥ 24 h, MDA含量显著大于胁迫6和12 h, 表明高浓度PEG和NaCl和长时间胁迫对花花柴叶片细胞造成很大伤害, MDA大量累积。
| 表1 PEG和NaCl胁迫对花花柴幼苗叶片MDA(μ mol· g-1)含量的影响 Table 1 Effect of PEG and NaCl stress on MDA(μ mol· g-1) content of Karelinia caspia leaves of seedling |
2.2.2 盐旱胁迫对花花柴幼苗叶片相对电导率的影响 细胞质膜是植物与外界环境相互作用的界面层, 是逆境胁迫对植物造成伤害的直接部位。不同胁迫时间和胁迫浓度对花花柴幼苗叶片相对电导率影响不同(表2)。在各处理条件下的叶片相对电导率均显著高于对照(P< 0.05)。当NaCl浓度≤ 300 mmol· L-1时, 胁迫12 h叶片相对电导率急剧升高到最高值, 胁迫24 h后趋于稳定; 而在400和500 mmol· L-1浓度下胁迫48 h时, 叶片相对电导率急剧升高到最高值, 在91.14%以上。在5%~20% PEG胁迫下, 花花柴叶片相对电导率随着胁迫时间增加呈先上升后降低的趋势, 并在胁迫12 h时达最大值, 12 h后趋于稳定。在25%胁迫下, 胁迫48 h时达最大值, 显著大于胁迫0、6和12 h。
| 表2 PEG和NaCl胁迫对花花柴幼苗叶片相对电导(%)率的影响 Table 2 Effect of PEG and NaCl stress on relative conductivity(%)of Karelinia caspia leaves of seedling |
2.2.3 盐旱胁迫对花花柴幼苗叶片SOD活性的影响 超氧化物歧化酶是植物体内清除活性氧的第一道防线。NaCl在200~400 mmol· L-1和PEG在5%~20%的胁迫下, 花花柴叶片SOD活性随着胁迫时间的延长而升高(表3); NaCl 500 mmol· L-1和25%PEG胁迫下的花花柴叶片SOD活性随着胁迫时间的增加呈先升高后降低的趋势, 并在胁迫12 h时达到最大值, 分别为1 065.64和1 008.40 U· (g· min)-1, 为对照的6.8和6.5倍。各NaCl和PEG胁迫处理下的叶片SOD活性显著高于对照(P< 0.05)。在200 mmol· L-1NaCl和5%PEG处理下, 花花柴幼苗在胁迫12~48 h内, 各处理间叶片SOD活性无显著差异(P> 0.05)。
| 表3 PEG和NaCl胁迫对花花柴幼苗叶片SOD[U· (g· min)-1]活性的影响 Table 3 Effect of PEG and NaCl stress on SOD[U· (g· min)-1] activity of Karelinia caspia leaves of seedling |
2.2.4 盐旱胁迫对花花柴幼苗叶片POD活性的影响 过氧化物酶可清除植物体内超氧化物歧化酶催化反应的产物过氧化氢, 从而植物细胞免受过氧化氢的毒害。各NaCl和PEG处理下的花花柴叶片POD活性显著高于对照(P< 0.05)(表4)。在NaCl 300 mmol· L-1和PEG 5%~20%处理下的花花柴叶片POD酶活性随着胁迫时间增加而增加; 在NaCl 200、400和500 mmol· L-1和PEG 25%胁迫处理下的花花柴叶片POD酶活性随着处理时间增加而呈先升高后降低趋势。叶片POD酶活性分别在NaCl 500 mmol· L-1处理12 h和PEG 25%处理24 h条件处理下达到最大值, 分别为3 536.59 U· (g· min)-1和3 480.59 U· (g· min)-1, 为对照的2.9和2.8倍。
| 表4 PEG和NaCl胁迫对花花柴幼苗叶片POD [U· (g· min)-1]活性的影响 Table 4 Effect of PEG and NaCl stress on POD [U· (g· min)-1] activity of Karelinia caspia leaves of seedling |
2.2.5 盐旱胁迫对花花柴幼苗叶片CAT活性的影响 过氧化氢酶也是植物清除活性酶系统的一种重要保护酶, 主要催化H2O2分解成H2O和O2。在NaCl溶液200~300 mmol· L-1和PEG 5%~20%胁迫下的花花柴叶片CAT酶活性随着胁迫时间增加而增加(表5); NaCl 500 mmol· L-1和PEG 25%胁迫处理下的花花柴叶片CAT酶活性随着处理时间增加呈先升高后降低趋势, 并分别在NaCl胁迫12 h和PEG胁迫24 h条时达到最大值, 分别为187.94和130.02 U· (g· min)-1, 为对照的6.3和4.4倍。各NaCl和PEG浓度处理下的花花柴叶片POD活性显著高于对照, NaCl 200~300 mmol· L-1处理6和12 h间花花柴叶片CAT酶活性无显著差异(P> 0.05), PEG 5%~20%处理24 h后, 各处理时间内的叶片CAT酶活性亦无显著差异。
| 表5 PEG和NaCl胁迫对花花柴幼苗叶片CAT [U· (g· min)-1]活性的影响 Table 5 Effect of PEG and NaCl stress on CAT [U· (g· min)-1] activity of Karelinia caspia leaves of seedling |
种子萌发是植物生长周期的重要环节, 水分和盐分是影响植物种子萌发的重要环境因子, 植物种子在萌发阶段的抗逆性可以反映该物种的抗逆性[9, 10]。对黑果枸杞(Lycium ruthenicum)[11]、罗布麻(A. venetum)[12]、沙芥(Pugionium cornutum)[13]等荒漠植物的研究表明, 低浓度盐旱胁迫对荒漠植物种子萌发无显著影响, 甚至会促进种子萌发; 而胁迫程度越高, 对种子萌发抑制越显著。在本研究中, 50 mmol· L-1的NaCl溶液和质量分数5%的PEG溶液处理的花花柴种子最终萌发率与对照无显著差异, 可能原因是种子在轻度渗透胁迫时, 具有补偿和超补偿效应[14], 低浓度的Na+、Cl-渗入种子, 降低种子渗透势, 加速吸水而促进种子萌发生长。而PEG和NaCl溶液浓度越高, 对花花柴种子渗透胁迫、离子胁迫伤害越大, 从而导致最终萌发率降低, 萌发进程滞后, 抑制作用越明显, 这与其他研究者对花花柴的研究结果一致[15]。
在正常生长环境下, 植物体内代谢过程中存在一个产生活性氧及清除活性氧的平衡, 因而植物不会累积活性氧。而在逆境下植物会产生大量的活性氧, 若不能及时消除, 则会引起细胞膜发生膜脂过氧化而受到伤害, 并产生丙二醛。因此, 丙二醛是衡量植物细胞膜受损害程度的重要指标。大量的研究表明, 植物叶片的丙二醛含量变化与其所受的逆境胁迫程度呈正相关, 胁迫程度越深, 细胞膜受损越严重, 丙二醛含量越大[16, 17]。本研究中, 在NaCl 200~300 mmol· L-1和PEG 5%~20%胁迫下, 花花柴叶片丙二醛含量随着胁迫时间的延长呈先上升后维持在稳定水平, 说明在轻度胁迫48 h内, 花花柴的膜稳定较强, 即使质膜受到损伤, 也会在一定时间后快速修复, 这也可能是其对逆境的一种重要适应机制。
在逆境胁迫下植株代谢紊乱, 诱导细胞内产生活性氧, 其与胞内生物大分子发生氧化反应, 生成具有强氧化性的膜质过氧化物和各种小分子的降解物, 导致膜质过氧化、破坏膜的完整性并降低保护酶的活性[18], 叶片相对电导率的变化是其最直观的反映。为了防御、减少氧化损害, 植物需要合理调节体内抗氧化清除系统, SOD、POD和CAT就属于这一系统中的关键酶。SOD作为重要的自由基清除酶, 能催化活性氧发生歧化作用而转化为H2O2和O2, 从而减轻超氧阴离子对植物体的毒害作用[19]。而SOD的歧化产物H2O2, 主要由CAT和POD协同清除, 共同防御活性氧大量积累造成的氧胁迫毒害[20]。本研究中, 各NaCl和PEG胁迫下的幼苗中3种酶的活性在前6 h内均迅速升高; 在12~24 h内, 3种酶活性分别逐步到达最高; 说明花花柴抗逆能力显著。400~500 mmol· L-1 NaCl和25% PEG胁迫处理下, 细胞受到严重的渗透胁迫和离子胁迫, 引起细胞内各代谢紊乱, 蛋白质变性, 保护酶失活, 清除ROS的能力降低, 最终导致细胞死亡。倪建中等[21]对木棉(Bombax ceiba)叶片的抗逆研究结果同样表明, 在重度胁迫下, 短时间内SOD活性迅速升高, POD和CAT活性逐步上升; 随着胁迫时间的延长, 这3种酶受到破坏, 活性逐渐降低, 叶片内ROS的积累超过其清理能力, 并最终造成木棉死亡。这与对万寿菊(Tagetes erecta)[22]和桑苗(Morus alba)[23]的盐胁迫特性研究结果一致。本研究表明, 在各浓度NaCl胁迫下, POD、SOD和CAT活性分别在24~48 h、12~24 h及12~24 h达到最大值; 在各浓度PEG胁迫下, POD、SOD和CAT活性在12~24 h达到最大值。根据酶活增长速率和到达峰值的时间不同, 对盐旱胁迫的敏感性SOD> CAT> POD。由此可见, SOD作为花花柴幼苗体内的第一道抗氧化防线, 能够迅速歧化多余的氧自由基, CAT和POD协同保护细胞免受伤害。
综上所述, 低浓度的盐旱胁迫对花花柴种子萌发和幼苗生理特性无显著影响, 花花柴种子萌发期和幼苗生长期的耐盐(NaCl)的临界值均为300 mmol· L-1; 耐旱(PEG-6000)临界值25%。花花柴种子萌发和幼苗生长期表现出较强的抗盐性和抗旱性, 这对荒漠区花花柴种群的建立和延续非常有利。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
盐旱胁迫对花花柴种子萌发与幼苗生理生化特性的影响
石新建
Physiological changes during seed germination and seedling development in Karelinia caspia Less. under drought and salinity stress
Shi Xin-jian