{kind=link}
{kind=link}
放牧强度对高寒草甸群落碳氮磷化学计量特征的影响
[李红琴1, 2 , 毛绍娟3 , 祝景彬1, 4 , 杨永胜1, 2 , 贺慧丹1, 4 , 李英年1, 2  ]
]
]
|
|


第一作者:李红琴(1981-),女,河南义马人,助理研究员,博士,主要从事草地生态系统碳循环研究。E-mail:hqli@nwipb.cas.cn
植被生态化学计量特征是高寒草甸生态系统稳定性的基础,但其对放牧管理的响应仍不清楚。为此,以2011年在青海海北高寒矮嵩草( Kobresia humilis)草甸建立的不同放牧强度试验地为研究对象,于2014年植物生长季的5月-9月,通过测定禁牧(对照,CK)、轻度放牧(light grazing,LG)、中度放牧(moderate grazing,MG)和重度放牧(heavy grazing,HG)样地的地上植被生产力和植被群落结构及群落叶片碳(carbon,C)、氮(nitrogen,N)、磷(phosphorus,P)含量,研究放牧强度对高寒草甸群落生态化学计量的影响。结果表明,MG能显著提高地上生物量和优良牧草禾草类植物的生物量比例,C含量的变异均在MG处理下最大,此结果支持了中度干扰理论;重牧能显著提高群落叶片的N含量( P<0.05);放牧管理对叶片P含量的影响无明显规律;禁牧有利于系统N和P的周转。本研究结果显示,适度的放牧强度有利于高寒草甸生产力的提高,而禁牧可能通过改变群落优势种增强系统的N、P循环强度。
Ecological stoichiometry characteristic is important for the stability of alpine meadow ecosystem; however, its response to grazing is still not clear. Grassland sample plots with different grazing intensity, including fencing (CK), light grazing (LG), moderate grazing (MG), and heavy grazing (HG), were established in the Haibei alpine meadow in 2011. These plots were chosen to investigate the effects of grazing intensity on aboveground vegetation productivity, carbon (C), nitrogen (N) and phosphorus (P) content of leaves of the vegetation community from May to September in 2014. The results showed that MG could significantly improve the aboveground productivity and proportion of Gramineae plants. Variation in the C content was maximum under MG treatment, supporting the hypothesis of intermediate disturbance. HG could significantly improve the N content. There was no significant effect of grazing on the P content. Fencing was beneficial to the turnover of N and P. The results indicated that moderate grazing was helpful for improving the productivity of the alpine meadow and fencing could enhance the cycle of N and P by changing the dominant species of the community.
生物的生长、发育以及行为离不开碳(carbon, C)、氮(nitrogen, N)、磷(phosphorus, P)3种元素。C构成植物体内干物质的50%左右, 是最主要的元素; N参与植物体内氨基酸、蛋白质、核酸等物质的生物合成, 并且在提高植物的光合作用等方面起着重要作用, 是植物体内的关键元素; P是植物生长发育不可缺少的营养元素之一, 它既是植物体内许多重要有机化合物的组分, 同时又以多种方式参与植物体内各种代谢过程[1]。化学计量内稳性是生物在长期的进化过程中适应环境变化的结果, 是生理和生化调节的反映。生态化学计量学是研究生物系统多种化学元素含量及其平衡关系的科学, 特别是C、N、P的计量关系[2]。植物叶片的C/N和C/P意味着生物量与养分的比值关系, 在一定程度上反映植物的养分利用效率[3]。
高寒草甸作为青藏高原草地生态系统的主要类型, 是当地畜牧业发展的主要基础[4]。然而, 近年来在气候变暖及人类活动的双重干扰下, 高寒草甸草地退化严重, 严重威胁到当地乃至我国的生态安全和人类生存与发展。放牧是草地生态系统最重要的利用方式之一, 过度放牧是人类施于草地生态系统最强大的影响因素, 全世界约有35%的草地退化是由过度放牧造成的[5]。放牧强度明显影响地上植物群落生产力、群落特征和地下营养水平[6, 7, 8], 势必影响草地生态系统物质循环和能量流动。有关放牧对草地土壤理化性质及养分的影响有较多报道, 普遍认为适度放牧是维护土壤养分及草地生产力的有效途径[9, 10, 11]。放牧管理对高寒草地植物生态化学计量的影响随着研究区域、土壤类型、植被类型以及放牧处理的不同而不同[12, 13]。本研究通过在青海海北同一牧户的草地, 建立不同放牧强度试验地, 探讨植物生长季不同阶段C、N、P元素含量和计量关系及对放牧强度的响应, 以期对放牧管理方式提出行之有效的措施和依据, 为平衡高寒草甸生态系统生产、生态功能提供理论依据。
试验在青海海北高寒草甸生态系统国家野外科学观测研究站(海北站)进行。海北站地处青藏高原东北隅祁连山北支冷龙岭东段南麓坡地(37° 29'-37° 45' N, 101° 12'-101° 23' E, 海拔3 200~3 600 m)。该区位于亚洲大陆腹地, 具有明显的高原大陆性气候, 东南季风微弱。冬半年受西伯利亚冷高压控制, 气候寒冷、干燥; 夏半年受微弱的东南季风影响, 气候凉爽、湿润。受高海拔条件制约, 气温极低, 无明显四季之分, 仅冷暖季之别, 干湿季分明。年平均气温-1.7 ℃, 降水量约560 mm, 降水主要集中于植物生长季的5月-9 月, 约占年降水量的80%, 年平均日照时数2 462.7 h[14]。植被建群种为矮嵩草(Kobresia humilis), 主要优势种为垂穗披碱草(Elymus nutans)、异针茅(Stipa aliena)、麻花艽(Gentiana straminea)、甘肃棘豆(Oxytropis kansuensis)和紫羊茅(Festuca rubra)等[15]。牧草生长低矮, 群落结构简单, 初级生产力低等。土壤为草毡寒冻雏形土(mat-cryic cambisols), 具有发育年轻、土层浅薄、有机质含量丰富等特征[16]。
冬季放牧强度试验平台于2011年8月建立, 参照以往研究放牧强度的经验, 牧压梯度设置为禁牧(对照, CK)、轻度放牧(LG, 4.5只羊· hm-2)、中度放牧(MG, 7.5只羊· hm-2)和重度放牧(HG, 15只羊· hm-2)4种管理方式。试验地用围栏围封, 试验羊为当地3~5龄成年藏系绵羊。放牧时间为9月20日到次年5月31日。
2014年5月-9月, 每月中旬左右, 以每个试验区中央点为中心, 设计20 m× 20 m的观测区, 以中央点及20 m× 20 m角点的正北方1 m处为观测样点, 即每个试验区共5个点(重复)。测定时用剪刀齐地面剪下25 cm× 25 cm样方内绿色植物装袋。置于105 ℃烘箱内杀青5 min, 然后置于60 ℃烘至恒重, 称重即为当月生物量。然后, 将样品研磨粉碎, 送至中国科学院西北高原生物研究所分析测试中心测定其C、N、P含量。于8月份分单种调查其生物量以了解其功能群的变化。
利用Excel 2010作图, SPSS 16.0进行统计分析。采用单因子方差分析(ANOVA)放牧强度对指标的影响, 采用最小显著差数法(LSD)比较各处理间的差异显著性。
CK、LG、MG和HG草地地上绿体生物量均表现为单峰型季节变化(表1)。在生长季初期(5月), CK草地的地上生物量最小, 随放牧强度的增大, 地上生物量依次增大。地上绿体生物量在8月份达到最大, MG最大, LG次之, HG和CK最小。9月份LG和MG生物量仍高于HG和CK。表明中度干扰有利于提高高寒草甸地上生产力。
| 表1 不同放牧强度高寒草甸地上绿体生物量季节动态 Table 1 Seasonal variation in aboveground biomass of the alpine meadow under different grazing intensities |
4个样地禾草类生物量比例大小顺序是MG> LG> CK> HG; 莎草类生物量比例大小顺序是HG> CK> MG> LG; 豆科类生物量比例大小顺序是HG> MG> LG> CK; 杂草类生物量比例大小顺序是CK> LG> HG> MG(图1)。牛羊喜食的禾本科所占比例表明, CK和HG均不利于高寒草甸生产性能的维持, MG最有利于高寒草甸禾本科牧草的生长。
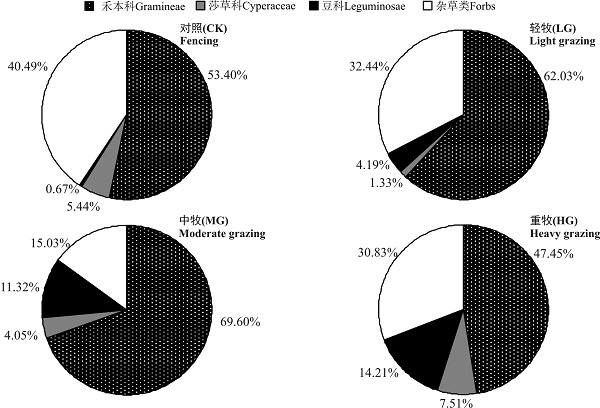 | 图1 不同放牧强度高寒草甸植物群落功能群生物量比例Fig. 1 Proportion of biomass of functional groups of the alpine meadow under different grazing intensities |
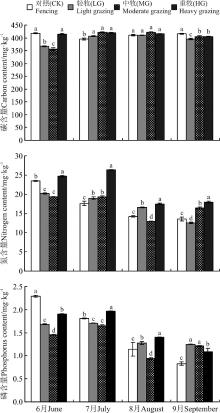
LG、MG草地群落叶片C含量在生长季初期的6月较低, 显著低于CK和HG(P< 0.05)(图2); 生长旺盛期, 放牧强度对群落叶片C含量的影响趋于减弱;
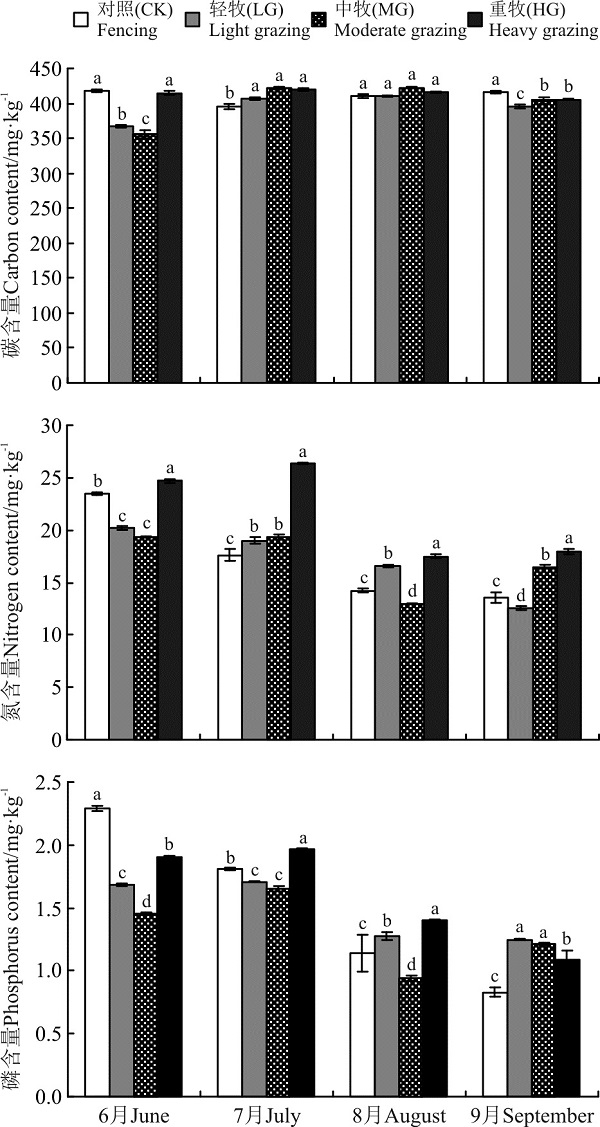 | 图2 不同放牧强度高寒草甸C、N、P含量的季节变化Fig. 2 Seasonal variation in C, N, and P content of the alpine meadow under different grazing intensities |
生长季末期9月, CK的C含量最大, 显著高于其余3个处理。以每个样地C含量的最小值为基准, CK、LG、MG和HG的C含量的变异系数分别是5.96%、12.07%、18.77%和3.55%, 表明中度放牧群落叶片C呈现出单峰型季节变化, 而CK和HG相对稳定。这表明, 中度放牧有利于系统C的周转。
HG群落叶片N含量显著高于其余3种处理, 表明重度放牧能显著提高群落叶片的N含量(P< 0.05), 而轻、中度放牧对N含量的影响无明显规律。另外, 群落叶片N含量随时间延长表现出逐渐降低的趋势。CK、LG、MG和HG的N含量的变异系数分别是72.96%、61.77%、49.27%和50.54%。表明禁牧和轻牧处理下植被群落N循环强度较高。
群落叶片P含量随时间也表现为逐渐下降的趋势(图2)。放牧降低了生长季初期的P含量, 而提高了生长季末期的P含量, 在生长季中期, LG、MG和HG对P含量的影响无明显规律。CK、LG、MG和HG的P含量的变异系数分别是175.90%、36.00%、75.53%和80.73%, 表明禁牧最利于群落的P周转。
一般线性模型分析(表2)表明, 放牧强度对群落叶片C、N、P含量的变化具有极显著的主效应(P< 0.01)。另外, 放牧强度的主效应对群落叶片的C、N、P含量的主效应远小于季节波动的主效应, 和二者的交互效应持平。
| 表2 放牧强度、季节动态下群落叶片C、N、P含量的一般线性模型 Table 2 GLM analysis of C, N, and P content between grazing intensity and month |
HG明显降低了植被的C/N, 其余3个处理之间差异不明显(表3)。放牧强度与P之间没有表现出一致的规律, 致使整个生长季C/P表现为MG> CK> HG> LG, 而N/P表现为HG> MG> CK> LG。
| 表3 不同放牧强度海北高寒草甸C、N、P化学计量特征 Table 3 Stoichiometry characteristics of C, N, and P in the Haibei alpine meadow under different grazing intensities |
放牧是人类对草地进行干扰的最主要方式, 它通过家畜的采食、践踏干扰以及排泄物的输入影响草地生态系统, 不仅改变草地的地上地下生物量、物种特征及群落组成, 还影响草地生态系统的能量流动和物质分配[17, 18, 19, 20]。本研究中, 5月份因植物刚返青, 生长缓慢, 地上生物量积累较少, 封育样地因枯落物太多, 阻碍植物生长, 放牧样地家畜的放牧采食使草地植物补偿性生长, 使得封育样地生物量低于放牧样地。6月-8月, 随着生长季雨热同期的来临, 草地植物生长迅速, 光合产物积累增加, 从而使草地地上活体生物量表现为增加趋势。放牧干扰下, 家畜的选择性采食以及践踏和粪便归还是植物群落类型转变的主要原因, 本研究表明, 中轻度放牧, 不仅能使草地地上净初级生产力(8月生物量)增大, 而且使家畜喜食的禾草类保持较高的比例, 与青海湖周边的高寒草原[21]研究结果一致。
对于同一植物种来说, 不受环境、发育时间影响的结构性物质, 其含量相对较稳定, 而功能性和贮藏性物质含量则受植物所处环境及生长发育阶段影响较大[22], 植物元素含量与自身结构特点和生长节律密切相关[23]。本研究通过对不同放牧强度海北高寒草甸生长季6月-9月植物群落采样分析发现, 群落叶片的C、N、P含量及其化学计量关系不但与放牧强度有关, 而且不同月份间也有明显差异。6月, 矮嵩草草甸大部分植物处于营养生长阶段, 植物体内功能性物质所占比例较大, 因而, 该月群落叶片N、P含量较高。随着生长季延长, 地上生物量进一步积累, 植物体内结构性物质所占比例增大, 因而, 7月和8月C含量最大。由于植物生物量的增加, 矮嵩草草甸植物体内N、P含量可能受到稀释效应的影响, 随生长季的延后逐渐减少。9月, 随着气温下降, 矮嵩草草甸植物停止营养生长, 转向生殖生长, 植物体内贮藏性物质向种子转移, 因而, 叶片N、P含量进一步减少。
有研究表明, 放牧对植物C、N、P含量影响主要通过以下两个方面实现:一方面放牧通过改变植物的形态和功能, 使C同化和积累的能力发生变化; 另一方面, 放牧后凋落物数量改变以及动物粪便可直接改变土壤营养状况; 同时, 动物践踏也使土壤理化性质发生改变, 影响根系对土壤C、N、P的吸收, 最终使植物的C、N、P含量及计量关系发生变化[24]。绵羊采食不仅刺激牧草生长, 还促使地上氮、磷向幼嫩器官重新分配, 从而使放牧草地地上活体的氮磷含量增加[25], 导致重牧样地生长季平均氮磷含量均最大。
豆科植物能与固氮菌结合成互惠共生体, 获得大量的N营养, 其氮含量及氮磷比高于其它功能群, 禾草可能因为有较高的营养利用率, 其N、P含量均显著低于其它功能群[26]。放牧能增加高氮低木质素即低碳含量物种数量, 从而使叶片C/N比降低[27]。本研究中, 重牧样地豆科植物比例大于其它3个样地, 因而其C/N最低, N/P最大。中度放牧样地, 禾草类植物比例较大, 因而其C/N和C/P较大。
植物营养吸收和C同化的途径不同, 一般情况下, C不会成为植物生长的限制因子, 而且C在植物体内含量很高, 变异较小, 因此, N和P的变化是影响C/N和C/P的主要因素[28]。本研究中, C的变异远小于N和P的变异, 与此结果一致。由于矮嵩草草甸植物生长迅速, N、P吸收与C的固定速率不成比例, C/N和C/P在9月份显著大于6月份, N/P变化较小。
MG能显著提高地上生物量和优良牧草禾草类植物的生物量比例, C的变异均在MG处理下最大, 支持了中度干扰理论。重牧能显著提高群落叶片的N含量, 放牧管理对叶片P含量的影响无显著规律。禁牧有利于系统N和P的周转。本研究结果显示, 适度的放牧强度有利于高寒草甸生产力的提高, 而禁牧可能通过改变群落优势种增强系统的N、P循环强度。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|