
{kind=link}
{kind=link}
{kind=link}
{kind=link}
梯度增温对青藏高原高寒草甸生态系统碳交换的影响
引用本文
耿晓东, 旭日. 梯度增温对青藏高原高寒草甸生态系统碳交换的影响. 草业科学, 2017,34(12):2407-2415
Geng Xiao-dong, Xuri. Ecosystem carbon exchange of an alpine meadow under simulated gradient warming in the Tibetan Plateau, China. Pratacultural Science,2017,34(12): 2407-2415
Permissions
Geng Xiao-dong, Xuri. Ecosystem carbon exchange of an alpine meadow under simulated gradient warming in the Tibetan Plateau, China. Pratacultural Science,2017,34(12): 2407-2415
Copyright©2017, 《草业科学》编辑部
梯度增温对青藏高原高寒草甸生态系统碳交换的影响
第一作者:耿晓东(1990-),男,安徽庐江人,在读硕士生,主要从事全球变化生态研究。E-mail:gengxiaodong@itpcas.ac.cn
摘要
高寒草甸是青藏高原主要的草地生态系统类型,对气候变化非常敏感,研究高寒草甸生态系统碳交换对升温的响应具有重要的理论和现实意义。在青藏高原中部地区的高寒草甸,使用开顶箱法(open-top chambers,OTCs)设置不增温对照(T0)以及4个不同程度的增温处理(T1、T2、T3、T4),采用CO2红外分析仪对生长季期间的碳交换进行连续3年的观测。结果表明,4个增温处理的5 cm土壤温度较之于不增温对照分别增加1.73 (T1)、1.83 (T2)、3.03 (T3)以及3.53 ℃ (T4);土壤水分没有发生梯度变化。观测期间,净生态系统碳交换(net ecosystem carbon exchange, NEE)基本为负值,因此高寒草甸表现为碳汇。增温小于2 ℃促进总生态系统生产力(gross ecosystem productivity, GEP),但对生态系统呼吸(ecosystem respiration, ER)影响较小,因而促进NEE,即促进高寒草甸的碳吸收;但增温大于3 ℃则抑制GEP,对ER影响较小,因而总体上对NEE产生抑制作用。综上所述,在高寒草甸生态系统,适度增温促进碳吸收,增温过度则降低碳吸收。
关键词:
增温; 高寒草甸; 碳交换; 呼吸; GEP; NEE; OTC
中图分类号:S812.29
文献标志码:A
文章编号:1001-0629(2017)12-2407-09
doi: 10.11829/j.issn.1001-0629.2017-0090
Ecosystem carbon exchange of an alpine meadow under simulated gradient warming in the Tibetan Plateau, China
Abstract
Alpine meadow is one of the primary alpine grassland ecosystems in the Tibetan Plateau region, which is sensitive to climatic changes, including global warming. Therefore, it is crucial to investigate how the ecosystem carbon exchange of the alpine meadow varies with increasing temperature. In the present study, we conducted a five-level warming experiment (T0, no warming; T1-T4 represent the different warming treatments) and the ecosystem carbon exchange was measured using open-top chambers (OTCs) and infrared analyzer, respectively, over three consecutive growing seasons. Our results showed that: 1) compared to the control, warming enhanced the temperature of 5 cm soil layer by 1.73 (T1), 1.83 (T2), 3.03 (T3) and 3.53 ℃ (T4), respectively for four warming treatments; but soil moisture changed in a non-linear fashion. 2) The negative values of the net ecosystem carbon exchange (NEE) indicated that the alpine meadow acted as a carbon sink. 3) Warming that less than 2 ℃ substantially improved the gross ecosystem productivity (GEP), but had no significant effect on ecosystem respiration (ER), resulting in a net increase in NEE value. In other words, moderate warming could stimulate carbon sequestration in the alpine meadows, but in a warmer scenario (e.g. temperature increased by more than 3 ℃), carbon sequestration would be restricted due to the decrease in GEP. These findings suggest that the alpine meadow will uptake more/less CO2 in moderate/outranged warming scenarios, accordingly.
Keyword:
warming; alpine meadow; carbon exchange; respiration; gross ecosystem productivity; net ecosystem carbon exchange; open-top chambers
全球气候变暖已是不争的事实, 由于温室效应, 预计到21世纪末全球气温将升高0.3~4.8 ℃[1]。青藏高原是气候变化的高敏感区, 其增温速率高于北半球同纬度地区[2], 近50年来的增温速率超过全球同期平均水平的2倍[3]。张人禾等[4]根据模型预测的结果, 预计到21世纪末, 青藏高原年平均气温将升高1.5~2.4 ℃, 且升温幅度存在地域和季节差异, 高原西南部升温强于东南部, 冬季升温高于夏季。
草地是最主要的陆地生态系统类型之一[5], 草地生态系统的稳定对维持区域以及全球性的生态系统平衡具有重要作用[6]。青藏高原是我国草地分布最广的区域之一[7], 独特的气候条件和高度的空间异质性孕育了多样化的高寒植被类型。其中, 高寒草甸是青藏高原最主要的植被类型之一, 约占青藏高原总面积的35%[8], 同时也是青藏高原重要的碳汇[9], 且具有很高的碳储量和释放潜力[10, 11]。研究表明, 增温可以通过直接和间接作用影响高寒草甸的碳交换[12], 但不同增温幅度对高寒草甸碳交换的影响还不明确。例如, Kato等[13]认为温度是控制净生态系统碳交换(net ecosystem carbon exchange, NEE)的主要因素; 但在不同条件下, 增温对NEE可能具有促进[14]或抑制[15]作用。通常认为增温促进高寒草甸总初级生产力(gross ecosystem productivity, GEP)[16], 但也有研究发现增温对GEP没有促进作用[17]。一些研究表明, 增温促进高寒草甸生态系统呼吸(ecosystem respiration, ER)[18, 19], 但也有研究认为其ER对温度变化敏感性不大[20]甚至受温度抑制[21]。有关三江源[22]和藏北[23]高寒草甸的研究表明, 增温条件下, GEP增加的幅度大于ER, 因而, 总体上增温促进生态系统的碳吸收(NEE)。
以上研究虽然都在高寒草甸生态系统进行, 但由于高寒草甸分布较广, 不同地区的本底温度并不一致, 且增温的幅度以及其他环境要素存在差异, 因此无法明确碳交换对不同幅度增温的响应。本研究依托中国科学院纳木措多圈层综合观测研究站, 在同一地点利用开顶箱法(open-top chambers, OTCs)设置4个增温梯度, 模拟不同的增温情景下高寒草甸碳交换的变化, 有助于准确认识高寒草甸生态系统应对气候变化的响应。
1 材料与方法
1.1 研究区概况
研究样地位于西藏当雄县念青唐古拉山北坡与纳木措湖东南岸之间(30° 44.08' N, 91° 1.06' E), 该地区海拔4 800~5 100 m, 为典型高寒草甸生态系统。2013-2014年生长季平均气温7.2 ℃, 2013年最高温度集中在6-8月, 最高气温13 ℃; 2014年最高温度集中在6-7月, 最高气温12.18 ℃; 2015年数据缺失。2013-2015年生长季平均降水量339 mm, 主要集中在7-9月。地上生物量为89.1 g· m-2, 土壤有机碳为29.1 mg· g-1, 土壤全氮为2 mg· g-1, 土壤pH为7.4[24]。植被分布以小嵩草(Kobresia pygmaea)为建群种。
1.2 试验方法
设置5个不同的开顶箱(OTCs)增温处理:不增温对照(T0)、增温处理一(T1)、增温处理二(T2)、增温处理三(T3)、增温处理四(T4), 每个处理设置3个重复。开顶箱方法为被动增温, 这在高海拔以及高纬度地区普遍应用[25]。这种装置为圆台状, 顶部开口直径为80 cm, T1、T2、T3、T4的装置底部直径分别为108.87、120.42、131.96、143.51 cm; 高度分别为25、35、45、55 cm。通过控制开顶箱的大小实现不同的增温幅度, 根据设计, 增温处理后, 各处理的相应温度关系为T0< T1< T2< T3< T4。在不同的处理区域埋设微型气象仪HOBO(H21, Onset Corp, Pocasset, USA), 记录地下5 cm处温湿度的季节动态变化; 使用雨量筒(RG-3, Onset Corp, Pocasset, USA)记录降水信息; 采用红外CO2分析仪(LI-840, LI-COR Inc., Lincoln, USA)观测不同处理碳交换的变化, 同时采用电子温度计(JM-624, 今明仪器有限公司, 天津)和手持湿度计(Delta-T Devices, Cambridge, UK)在观测时同步测量地下5 cm土壤温度和湿度状况。
NEE/ER观测:在增温处理样地中心将边长50 cm的正方形底座嵌入土壤, 观测时将边长50 cm的正方体透明塑料箱(5面, 内置小型风扇)罩在底座上, 形成一个相对密闭的空间, 并通过空气导管连接红外分析仪, 并同时启动箱内风扇使得空气混合均匀。分析仪每秒记录一次箱内CO2浓度, 记录约120 s即完成一次观测。使用透明塑料箱时, 箱内同时存在光合与呼吸作用, 此时记录的为NEE; 当使用遮光布套住塑料箱, 阻断光照, 箱内只有呼吸作用, 此时记录的为ER。根据Jasoni等[26], NEE和ER计算公式为:
F=
式中:F表示NEE和ER的通量, NEE为负值表示生态系统吸收CO2; V表示箱内体积(m3); P表示观测时箱内平均大气压(kPa); W表示观测时箱内平均水汽分压(mmol· mol-1); R表示标准气体常数[8.314 J· (mol· K)-1)]; S表示底座面积(m2); T表示观测期间箱内平均气温(℃); C'表示经过水汽校正的CO2浓度, C'=C/(1-W/1 000), C表示红外分析仪观测时记录的箱内CO2浓度(μ mol· mol-1); dC'/dt表示箱内CO2浓度随时间变化的速率。GEP的计算公式为:
GEP=-NEE+ER。
OTC装置在观测前两个月架设。2013年架设于6月份并在8月21日进行3个重复的观测, 2014年架设于5月份并在7月24日、8月3日、8月19日和9月11日进行了10个重复的观测, 2015年架设于7月份并在8月9日、8月12日、9月4日和9月13日进行了5个重复的观测。观测时间为晴朗天气的09:00左右。
1.3 数据处理
采用非参数检验法Tamhane’ s T2检验不同处理之间的土壤温度、水分之间的差异, 采用邓肯新复极差法(Duncan)多重比较不同处理间NEE、ER以及GEP的差异; 皮尔逊(Pearson)相关和简单回归关系法分析碳交换通量与温度的关系。统计分析使用SPSS 22.0完成, 插图使用Origin 9.1 绘制。
2 研究结果
2.1 处理样地的温度与水分变化
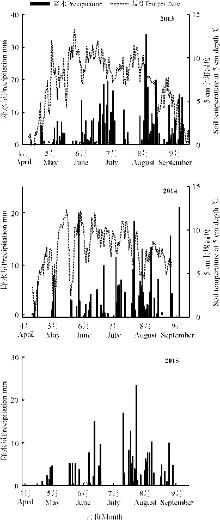
2013年生长季累积降水量486.2 mm, 主要集中在7-9月, 但8月份降水主要集中在下半月; 2014年生长季降水分布较为均匀, 累积降水量285.6 mm, 最高降水为8月份; 2015年累积降水量245.1 mm, 主要集中在6月和8月, 7月下旬到8月初没有降水, 维持了一段干旱时期, 土壤温度的变化与降水趋势基本一致(图1)。
增温处理后, HOBO记录了2013年的5-9月、2014年的7-9月的5 cm土壤温湿度; 2015年, 因设备损坏, 在8-9月采用电子温度计以及手持湿度计补测土壤温湿度。结果表明, 相对于对照T0, 增温处理T1、T2、T3、T4的土壤温度基本上依次升高。2013年, 分别增温2.0、1.9、2.5、3.1 ℃; 2014年, 分别增温1.2、1.4、2.8、3.6 ℃; 2015年, 分别增温2、2.2、3.8、3.9 ℃。根据HOBO观测的数据, 2013和2014年不同处理间土壤水分出现明显差异, 但水分变化趋势与增温梯度并不一致; 2015年通过湿度计观测, 各处理土壤水分没有发生显著变化(图2)。
2.2 不同增温处理之间碳交换的变化
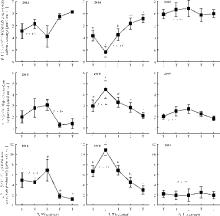
2013年, 不同处理间的NEE、ER没有显著差异(P> 0.05), T2处理的GEP、ER以及NEE具有最大通量, 分别达到6.85、3.06和-3.79 μ mol· (m2· s)-1, T4处理的GEP显著低于T1处理。2014年, 不同处理间的NEE、ER以及GEP均具有显著差异(P< 0.05), T1处理的GEP、ER以及NEE具有最大通量, 分别达到10.84、4.48和-6.35 μ mol· (m2· s)-1, 较之于T0以及T2, T1处理的GEP和NEE通量显著升高, 而ER变化不显著(P> 0.05)。2015年没有观测到NEE、ER以及GEP在各不同处理之间存在显著差异(图3)。
2.3 温度与碳交换的关系
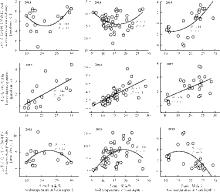
NEE、GEP与土壤温度呈二次相关关系(2013年这种关系不显著), 温度较低时, 增温促进NEE和GEP通量, 例如, 2013和2014年土壤温度低于20 ℃时, 增温对NEE和GEP的通量具有促进作用; 当土壤温度高于20 ℃时, 增温则具有抑制作用(图4)。2015年, 当温度高于15 ℃时, 增温降低NEE和GEP的通量具值。ER与土壤温度呈正相关关系, 即随着温度增加, 生态系统呼吸显著加快(P< 0.05)。
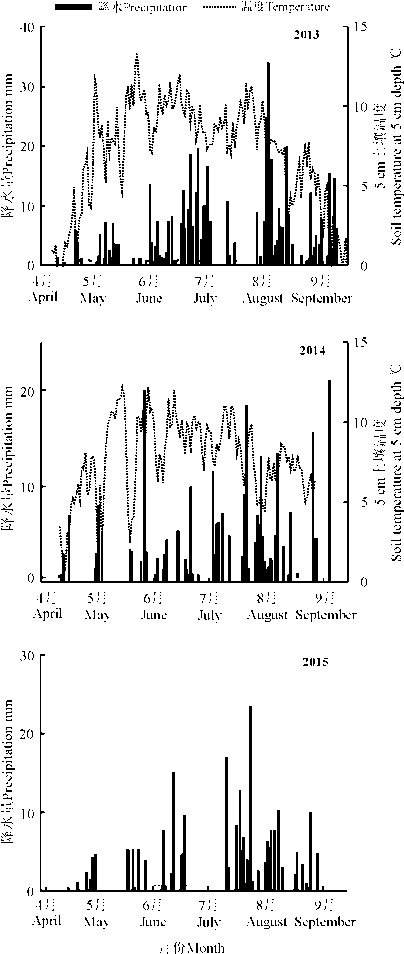 | 图1 高寒草甸生长季降水量和5 cm土壤温度动态变化 注: 2015年缺失土壤温度动态数据。Fig. 1 Seasonal variation in precipitation and soil temperature at 5 cm depth in the alpine meadow Note:The dynamic data of soil temperature was not available in 2015. |
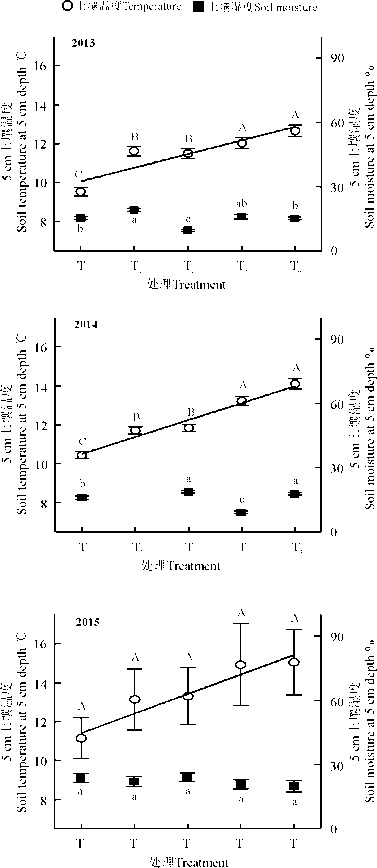 | 图2 不同增温处理下5 cm土壤温度和湿度 注:T0-T4表示不同增温处理, 不同字母表示増温处理间差异显著(P< 0.05), 图3同。其中, 大写字母表示处理间土壤温度差异; 小写字母表示处理间土壤湿度差异情况。Fig. 2 Soil temperature and moisture in 5 cm soil for different warming treatments Note:T0-T4 represent different warming treatments, different letters indicate significant difference among different warming treatments at the 0.05 level, similarly for the Fig.3. Specially, the uppercase and lowercase letters reflect the differences of soil temperature and moisture among different treatments. |
3 讨论
3.1 模拟增温对GEP的影响
已有研究表明, 增温(1~2 ℃)能显著促进高寒草甸生态系统的GEP[23, 27]。这与本研究2014年T1增温处理的研究结果一致; 但当增温的幅度更大时, 则出现了不同的结果, 例如, 2014年, T4处理增温3.6 ℃, GEP显著下降(图3)。这是因为在高寒草甸, 增温会改变植被特征以及物种光合特性等[28], 当土壤温度超过20 ℃, GEP通量与温度呈负相关关系(图4), 从而导致GEP降低。另外, 在高寒草甸生态系统, 温度对碳通量的影响, 常常受到土壤水分的制约[22]。例如, 有研究表明, 只有在土壤水分不是制约因子时, 增温才提高了植物生产力[29, 30]。因此当增温幅度过大时, 可以间接通过土壤水分作用导致GEP降低, 这是在土壤温度较高时, 增温对GEP产生影响的另一个方式。2013年的观测频率较低, 因而在统计学上出现偶然的几率偏大, 较大幅度的增温处理(如T3和T4), 其GEP、ER以及NEE的通量偏低, 可能是高温影响了地上植物的生长所致。2015年OTC装置的安装较迟, 导致随后的观测数据没有体现出增温效果(图2、3)。
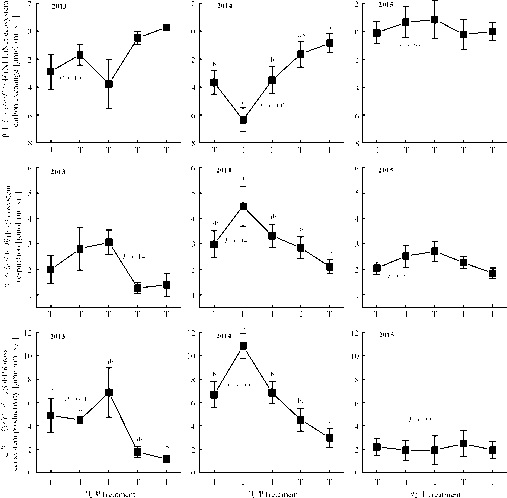 | 图3 不同增温处理净生态系统碳交换(NEE)、生态系统呼吸(ER)以及总生态系统生产力(GEP)Fig. 3 Seasonal average net ecosystem carbon exchange, ecosystem respiration and gross ecosystem productivity for different warming treatments |
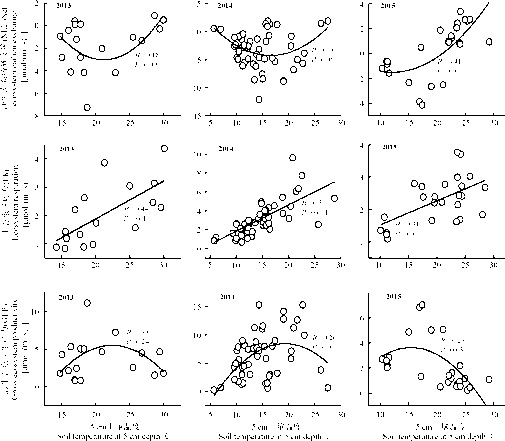 | 图4 净生态系统碳交换(NEE)、生态系统呼吸(ER)以及总生态系统生产力(GEP)分别与5 cm土壤温度的相关关系Fig. 4 Dependency of net ecosystem carbon exchange, ecosystem respiration and gross ecosystem productivity on soil temperature in the alpine meadow |
3.2 模拟增温对ER的影响
生态系统呼吸(ER)包括植物呼吸和土壤呼吸两个过程。首先, 增温显著提高地上植被呼吸强度[31]; 其次, 增温促进土壤相关酶活性[32]以及微生物的生理活动[33]。因此, 总体上在适度的温度范围内, ER与土壤温度具有正相关关系(图4)。就不同增温处理而言, 除了温度的改变, 土壤水分也发生变化, 土壤水分在调节土壤透气性以及土壤基质溶解性扩散等方面具有重要作用[34], 也与部分土壤酶的活性显著相关[35]。增温造成的土壤水分变化(图2), 直接影响到土壤微生物对基质的获取能力, 从而影响土壤呼吸的温度敏感性[36]。由于土壤水分的变化, 增温可能对土壤呼吸具有抑制作用。例如, 有研究发现增温在增加植物呼吸的同时, 也降低了土壤呼吸[31]。在高寒草甸, 土壤呼吸占据生态系统呼吸的重要比例(约46%)[20]。因此, 各不同增温处理的ER在生长季水平上基本没有显著的变化(2014年, T3和T4处理的生态系统呼吸较低是由于增温幅度过大影响了植物的生长) (图3), 可能是由于土壤呼吸受到较强的抑制所致。
3.3 模拟增温对NEE的影响
增温促进土壤呼吸、降低土壤有机碳含量, 同时也增加来自植物的碳输入, 因此在不同的条件下, 增温可能引起NEE的增加、降低或者不变, 导致生态系统可能表现为碳汇, 也可能表现为碳源[37]。在本研究的增温处理中, NEE总体上为负值, 说明高寒草甸生态系统表现为碳汇(图3)。增温对NEE的影响取决于GEP和ER的变化, 如图3所示, 增温对GEP的影响超过对ER的影响, 因而NEE的变化更多取决于GEP对增温的响应。2013和2015年的观测数据存在缺陷; 2014年的结果呈现两个不同的变化趋势:T1处理GEP较未增温对照显著增加, 而ER的变化不显著, 因而直接导致了NEE的显著增加(即增温T1 处总体上促进对碳的吸收); 当增温幅度偏大, 如T4处理, 则导致了GEP显著降低, ER没有显著变化, 因而最终导致NEE通量降低(图3)。这主要源于在高寒草甸, 当土壤温度较低时, 增温促进NEE通量, 而当土壤温度较高时, 增温则对NEE通量产生抑制(图4)。说明在高寒生态系统, 如果继续增温, 会造成地上生产力的降低和土壤呼吸的增强[38, 39], 并最终可能导致碳排放[40]。多个生态系统的研究发现增温显著促进植物光合以及生态系统呼吸, 并最终促进碳吸收[41], 本研究结果表明, 在高寒草甸生态系统, 适度增温可以促进CO2的吸收, 但过度增温会导致碳吸收降低。
4 结论
目前, 高寒草甸在生长季表现为碳汇。在高寒草甸生态系统, 增温对GEP的影响大于对ER的影响; 适度增温(如低于2 ℃)促进NEE通量, 即增强生态系统的碳吸收, 但过度增温则会降低生态系统的碳吸收。
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|