

{kind=link}
{kind=link}
{kind=link}
{kind=link}
利用EST-SSR标记构建中国老芒麦品种DNA指纹图谱及种质遗传多样性
[张俊超, 谢文刚 , 赵旭红, 张宗瑜, 赵永强, 王彦荣]
, 赵旭红, 张宗瑜, 赵永强, 王彦荣]
, 赵旭红, 张宗瑜, 赵永强, 王彦荣]
|
|


王彦荣(1956-),女,吉林大安人,教授,博士,主要从事草类种子及种质资源研究。E-mail:yrwang@lzu.edu.cn
第一作者:张俊超(1990-),男,河南驻马店人,在读博士生,主要从事牧草种质资源与育种研究。E-mail:zhangjch16@lzu.edu.cn
本研究利用EST-SSR分子标记,对国内的52份老芒麦( Elymus sibiricus)材料进行遗传多样性的研究,并构建了7个老芒麦品种及栽培材料的DNA指纹图谱。20对EST-SSR引物共产生204个条带,平均每对引物扩增出10.2条带,176(86.27%)条带为多态性条带,表明供试材料具有较高水平的多态性。同时,引物Elw404和Elw195构建的指纹图谱较为容易地将品种和栽培材料与其他材料区别开。分子方差分析(AMOVA)表明,老芒麦地理区域内(73.94%)的遗传变异远高于地理区域间(26.06%)。结构分析将52份材料分为5组,主成分分析(PCoA)结果与之相似。与品种及栽培材料相比,野生材料具有更高的遗传多样性[多态性条带数(NPB)为174,多态性百分比(PPB)为85.29%,香农多样性信息指数(I)为0.296 6,Nei’s基因多样性(H)为0.179 8,观察等位基因数(Na)为1.852 9],可作为重要的遗传资源用于后续的育种工作。本研究结果表明,EST-SSR分子标记可以有效地评价老芒麦种质遗传多样性及进行品种鉴定。研究结果为制定老芒麦种质原位和非原位保护策略提供参考。
In this study, EST-SSR markers were used to analyze the genetic diversity of 52 Elymus sibiricus accessions, and to construct DNA fingerprinting profiles of seven E. sibiricus cultivated accessions and cultivars in China. A total of 204 bands were generated from 20 EST-SSR markers with an average of 10.2 bands per marker. Of the total, 176 (86.27%) bands were polymorphic, indicating that the tested materials possessed a high level of polymorphism. The primers Elw404 and Elw195 could distinguish the seven cultivated accessions and cultivarsfrom other wild materials. Analysis of molecular variance showed that genetic variation was greater within geographical regions (73.94%) than between them (26.06%). Structure analysis suggested that the 52 accessions were clustered into five groups; a similar result was obtained using principal coordinate analysis. Compared with cultivars and cultivated accessions,wild accessions[number of polymorphic bands=174, percentage of polymorphic bands (PPB)=85.29%, Shannon’s information index (I)=0.296 6, Nei’s gene diversity (H)=0.179 8, observed number of alleles (Na)=1.852 9] possessed higher genetic diversity, suggesting that wild accessions could be used as important genetic resources for future breeding programs. Furthermore, ex situ and in situ conservation strategies were suggested for E. sibiricus germplasm. The results of the present study showed that EST-SSR markers provided an efficient means of assessing genetic diversity and identifying cultivars in E. sibiricus.
准确地鉴别植物栽培材料、品种及野生材料, 并明确它们的遗传关系, 是育种研究、品种管控与登记, 以及保护育种者权利的基础[1]。传统方法中, 植物品种的鉴别常使用形态学与生育期特性相结合的方法, 但这种方法具有比较困难、定义不明确、耗时且具主观性的缺点。同时, 鉴定的准确性会受环境因素的影响而受限制。DNA指纹图谱技术则可有效地弥补传统方法的缺陷, 这种新方法可以将植物任何生理时期的任何组织作为材料。很多分子标记, 诸如RAPD、ISSR、SSR、SRAP、SCOT[2, 3, 4, 5, 6]等, 已经成功地在许多作物和牧草物种中用于品种鉴别和遗传多样性的测定。其中, SSR标记具有高突变率、共显性和高重复性的优点, 可作为理想的标记方法, 用于牧草及一些农业和园艺植物的品种鉴定和遗传多样性的研究[7, 8, 9, 10, 11]。
披碱草属(Elymus)是小麦族(Triticeae)中最大的属, 它包含遍及世界的150个物种[12]。老芒麦(E. sibiricus), 又称西伯利亚野黑麦(Siberian wild rye), 在世界范围分布广泛, 如在东欧、西伯利亚、中亚、西亚、蒙古、朝鲜、及印度均有分布[13], 在北美洲也有少量分布, 主要集中在亚北极地区, 即阿拉斯加和加拿大北部[14, 15]。国内老芒麦的种质丰富且多样化, 它的主要分布地区包括河北、内蒙古、山西、陕西、西藏、青海、四川、甘肃及新疆等省(区)[4, 5, 6, 16]。老芒麦是一种多年生、自花授粉的异源四倍体牧草[14]。它具有对环境适应性强, 耐寒、耐旱性良好以及出色的草品质, 已广泛应用于天然草地和栽培草地。但老芒麦在品种选育和遗传改良方面的进展较为缓慢。到目前为止, 国内仅登记了8个老芒麦品种, 其中的一些品种有着相对狭窄的遗传背景。这可能与育种的原始材料在自然或人工条件下与已有推广品种发生杂交有关[17]。因此, 老芒麦品种选育进展缓慢的原因之一是缺少有效的遗传资源和遗传变异。鄢家俊等[4]采用SRAP和SSR分子标记研究了青藏高原地区的野生老芒麦种质遗传多样性, 结果表明该地区的老芒麦种质具有丰富的遗传多样性。Xie等[18]基于EST-SSR标记研究发现, 来自青藏高原地区具有不同落粒性的老芒麦种质的遗传多样性水平较高。Xie等[6]采用ScoT分子标记, Ma等[19]运用醇溶蛋白标记, 均发现世界来源的老芒麦材料具有高水平的遗传变异, 且遗传分化与其生态地理环境有密切关系。然而, 针对国内老芒麦主要分布地区的种质遗传多样性和遗传关系并未有较好的论述。构建老芒麦品种和栽培材料的DNA指纹图谱, 在国内外报道较少[17, 20]。因此, 借助EST-SSR分子标记构建国内老芒麦品种指纹图谱, 并对国内野生老芒麦材料和品种间进行深层次的遗传多样性比较分析, 可以对老芒麦遗传多样性分布提供新的见解。
本研究利用EST-SSR分子标记构建7个老芒麦品种及栽培材料的DNA指纹图谱, 评价与比较品种、栽培材料和野生材料间的遗传多样性和亲缘关系, 以期促进老芒麦种质收集、保护和育种工作。
研究所用的52份老芒麦材料均来自国内, 主要包括品种、栽培材料和野生材料(表1)。其中包括4个栽培品种(同德、青牧1号、川草2号、红原), 18份引种材料由美国农业部提供, 其他材料由兰州大学草地农业生态系统国家重点实验室, 四川农业大学和四川省草原科学研究院提供。所有的材料按起源地和自然区划可归结为5个地理区域, 分别为青海(QH)、新疆(XJ)、四川(SC)、内蒙古(NM)和甘肃(GS)。
| 表1 试验中老芒麦材料的信息 Table 1 Elymus sibiricus accessions used in the study |
每份材料选取25个幼苗的叶片等量混合, 经冻干处理后采用改进的CTAB法提取DNA[21]。通过使用Nanodrop分光光度计和琼脂糖凝胶电泳对DNA的浓度和质量进行测定。最后, DNA的浓度使用ddH2O调整为25 ng· μ L-1用以进一步的分析。
本研究中, 100对EST-SSR引物主要来源于披碱草属、拟鹅观草属(Pseudoroegneria)及赖草属(Leymus)[22, 23]。最终, 筛选出20对多态性高的EST-SSR引物用于52份老芒麦材料遗传多样性评价。PCR扩增反应在PTC-200热循环仪上进行, 扩增程序:94 ℃变性15 s, 65→ 60 ℃退火30 s, 72 ℃延伸1 min, 共5个循环; 接下来是94 ℃变性30 s, 60 ℃退火30 s, 72 ℃延伸1min, 共30个循环; 最后72 ℃延伸7 min; 4 ℃保存。扩增片段点样于6%变性聚丙烯酰胺凝胶, 电泳后, 凝胶用AgNO3溶液染色, 显影后在凝胶成像系统拍照。
对清晰度高、分辨度好的扩增条带按照条带的有无分别记为1, 0。参照以下公式[24]计算每一对EST-SSR引物的多态信息含量值(PIC):
PIC=1-p2-q2.
式中:p表示有带所占的频率, q表示无带所占的频率。条带的信息量(Ib)按照如下公式[25]计算:
Ib=1-(2× |0.5-p|).
式中:p表示有带的品种或基因型所占的比例。
条带的分辨率(Rp)通过如下公式来计算[24]:
Rp=∑ Ib.
POPGENE 32 v1.31软件[26]用在假定Hardy-Weinberg平衡条件下计算这些材料的遗传一致度(GI)、遗传距离(GD)、多态性条带数(NPB)、多态性百分比(PPB)、香农多样性信息指数(I)、Nei’ s基因多样性(H)及观察等位基因数(Na)。采用AMOVA v1.55软件分析类群内和类群间遗传变异[27]。STRUCTURE v2.3.4软件[28, 29]进行遗传结构分析。由NTSYS v2.10软件进行主坐标分析, 同时利用品种和栽培材料的数据, 结合Winboot软件构建系统树图[30, 31]。其中, POPGENE和AMOVA的输入文件由DCFA v1.1软件准备[32]。
本研究中, 20对EST-SSR引物是由100对EST-SSR引物筛选而来, 这些引物可扩增出多态性高且清晰的条带(表2)。每对引物扩增出的多态性条带数量从2(Ps3577)到21(Elw306)不等, 平均值为8.8。20对EST-SSR引物的多态信息含量(PIC)变化范围为0.26(Ps3577)~0.50(Ps1475, Elw228)。条带分辨率(Rp)从0.27(Ltc0055)到7.42(Elw404)不等, 平均值为3.01。基于以上结果, Elw306(总条带数和多态性条带数均为最大值)、Elw404(Rp为最大值)、Ps2283、Elw195和Elw187这5对引物在遗传多样性分析中具有更多的潜力, 可以较少的引物研究更多的个体和取样点。
| 表2 20对EST-SSR引物用于老芒麦品种、栽培材料和野生材料基因型的研究 Table 2 20 EST-SSR primers used to genotype Elymus sibiricus cultivars and, cultivated and wild accessions |
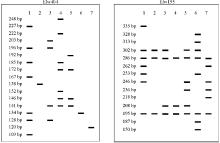
引物Elw404和Elw195的条带分辨率及多态信息含量值较大, 被选作用于构建7份老芒麦品种和栽培材料的DNA指纹图谱(图1)。两对引物扩增的条带数分别为17和14, 条带大小范围为109~335 bp, 这些独一无二的指纹图谱很容易将7份供试材料相互区别开。
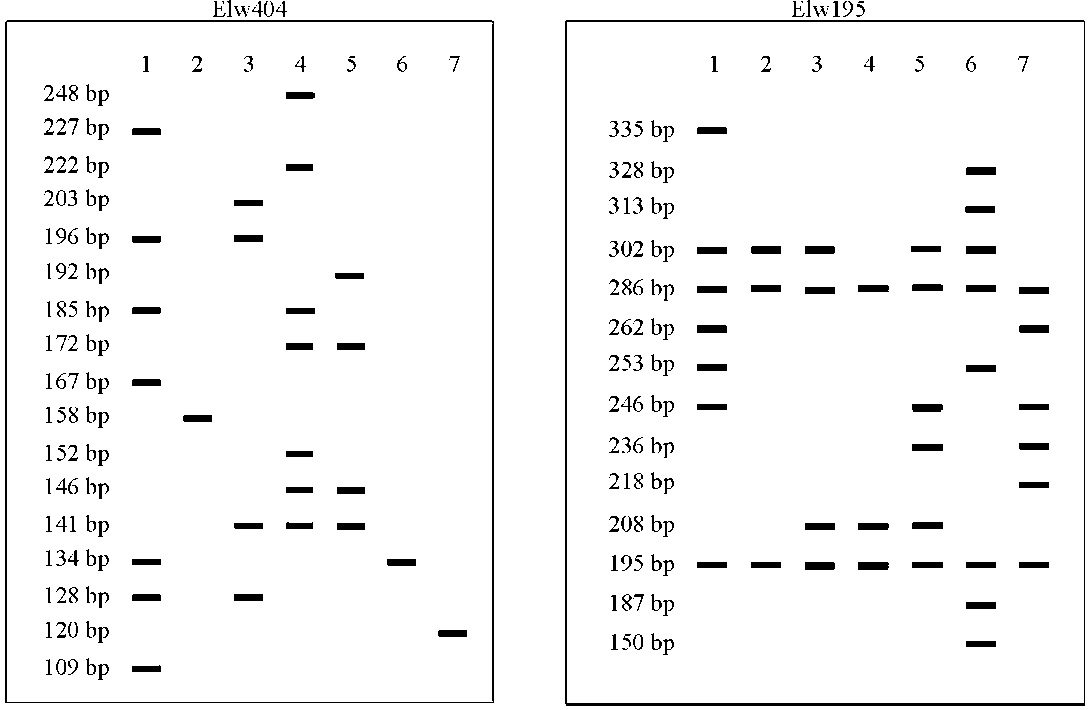 | 图1 通过Elw404和Elw195两对引物构建老芒麦品种(栽培种)指纹图谱 注:1号材料为同德, 2号材料为青牧1号, 3号材料为PI499468, 4号材料为川草2号, 5号材料为红原, 6号材料为PI499457, 7号材料为PI499456。Fig. 1 Molecular fingerprinting of Elymus sibiricus cultivars (cultivated accessions) using the primers Elw404 and Elw195 Note: 1, Tongde; 2, Qingmu No.1; 3, PI499468; 4, Chuancao No.2; 5, Hongyuan; 6, PI499457; 7, PI499456. |
20对引物共扩增出204条带, 其中176条带(86.27%)为多态性条带(表1), 表明52份老芒麦材料具有高水平的遗传多态性。此外, 与品种和栽培材料(PPB=50.98%, I=0.234 7, H=0.151 0, Na=1.509 8)相比, 野生老芒麦材料具有更大的变异性(PPB=85.29%, I=0.296 6, H=0.179 8, Na=1.852 9)(表3), 这一结果可能与试验中栽培材料数量较少有关。同时表明野生老芒麦材料有着广泛的遗传背景, 可作为重要的基因库用于品种改良。
| 表3 EST-SSR标记用于老芒麦品种和栽培材料与野生材料的遗传变异性 Table 3 Genetic variability between cultivar & cultivated accessions and wild groups of Elymus sibiricus detected by EST-SSR markers |
AMOVA分析表明, 老芒麦地理类群内部的遗传变异为73.94%, 地理类群间的遗传变异为26.06%(表4)。5个地区材料的遗传一致度(GI)从0.941 0到0.895 2不等(表5)。青海和新疆材料间的遗传一致度最小, 为0.895 2。四川与甘肃材料间的遗传一致度最大, 为0.941 0, 两个类群材料均采样于青藏高原的东北缘。
| 表4 基于EST-SSR标记对老芒麦的分子方差分析 Table 4 Analysis of molecular variance of Elymus sibiricus based on EST-SSR markers |
| 表5 老芒麦5个地理类群间Nei’ s遗传一致度(对角线上部)和遗传距离(对角线下部) Table 5 Pairwise Nei’ s genetic identity (above diagonal) and genetic distance (below diagonal) for five geographical populations of Elymus sibiricus |
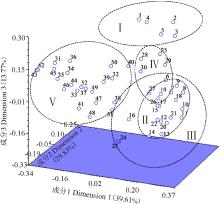
借助STRUCRURE V 2.3.4软件, 基于哈迪温伯格(Hardy-Weinberg)平衡, 对52份材料的遗传结构进行评估, 根据最大似然值和Δ K值可得到类群数目的最佳值(图2)。K=5时, 所有的材料聚集为5个类群, 这与老芒麦地理分布几乎一致, 因此5就是最合适的类群数目。类群Ⅰ 包括来自青海省的2个品种和3份野生材料; 类群Ⅱ 包含来自新疆的1个栽培材料和9份野生材料; 类群Ⅲ 包括来自四川省的2个品种和7份野生材料; 类群Ⅳ 包含来自内蒙古的2个栽培材料和4份野生材料; 最后一个类群包括来自甘肃省的22份野生材料。主坐标分析遗传关系的结果与结构分析一致:前3个主成分可解释69.44%的总变异, 它们分别占39.61%、29.83%和13.77%(图3)。
 | 图2 中国52份老芒麦材料遗传结构分析 注:类群编码同表1所示, 不同的个体具有相同的颜色表示它们归属为同一类群。Fig. 2 STRUCTURE analysis of 52 Elymus sibiricus accessions in China Note: The population codes are indicated in Table 1. Same color in different individuals indicates that they belong to the same cluster. |
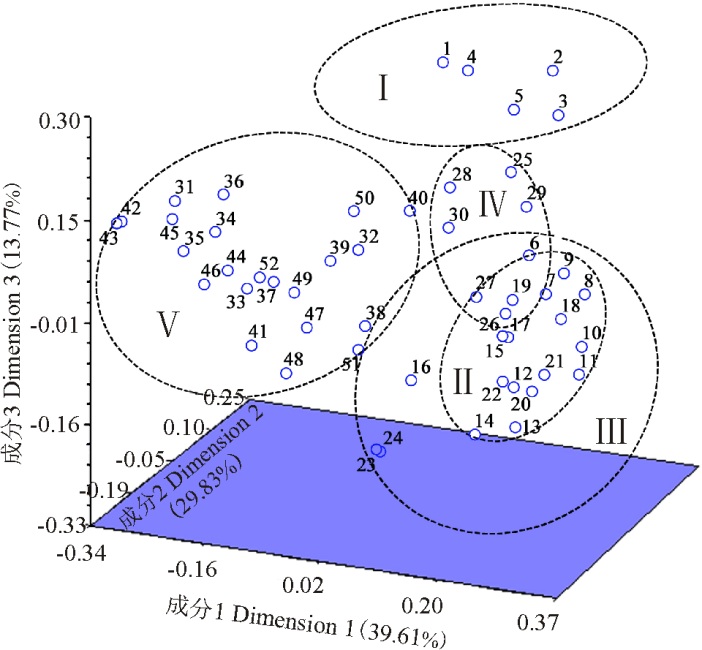 | 图3 基于EST-SSR标记使用Dice遗传相似矩阵对52份老芒麦进行主坐标分析的三维发散图Fig. 3 Principal coordinates analysis of three-dimensional scatter diagram for EST-SSR markers using the Dice genetic similarity matrix for 52 Elymus sibiricus accessions |
利用统计所得数据, 将3个品种和4份栽培材料单独进行聚类, 由此得到的系统树图揭示了这几份材料间的遗传关系(图4)。这些材料聚为两大类(Ⅰ 和Ⅱ ), 并且类群Ⅰ 包括3个亚类群(A, B, C)。3个省(区)的6份材料聚在类群Ⅰ , 同一地区的2份材料又形成单独的亚类群, 而类群Ⅱ 仅包括来自新疆的1个栽培材料。
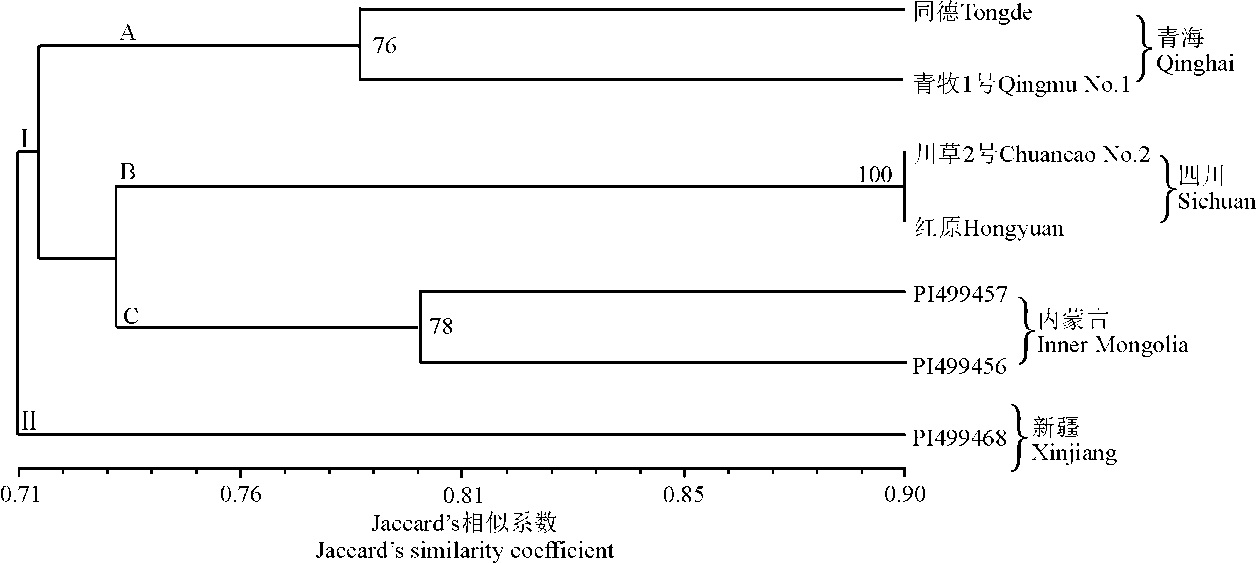 | 图4 基于Jaccard’ s遗传相似性及非加权组平均法推导出老芒麦品种和栽培材料的系统树图 注:图中只表示出bootstrap值大于50%的部分。Fig. 4 UPGMA-derived dendrogram of Elymus sibiricus cultivar and cultivated accessions based on Jaccard’ s genetic similarity Note: Only bootstrap values higher than 50% are presented. |
老芒麦种质遗传多样性的研究, 既可促进对该物种种质资源的管理、评价及利用, 也是创制老芒麦优异新品种的基础工作[35]。本研究中, 利用EST-ISSR引物扩增的多态性条带百分比(PPB)为86.27%, 表明老芒麦具有高水平的遗传多样性。该结果与前人的研究结果相似, 如SRAP(PPB=86.48%)[4], SSR(PPB=86.88%)[5], 高于ISSR(PPB=77.20%)[3]和RAPD(PPB=78.65%)[2], 但是低于SCoT(PPB=89.0%)[6]和醇溶蛋白(PPB=90.4%)[19]。老芒麦高水平的遗传多样性可能是由于其地理来源多样化和分子标记的高效性, 在象草(Pennisetum purpureum)[36]和大豆(Glycine max)[37]的研究报道中已证明地理来源和遗传多样性之间有较强的相关性, 严学兵等[38]认为地理生态环境与披碱草属植物的遗传变异有关, 并指出青藏高原与蒙古高原多变的气候与环境因素会引起该属同种不同居群间遗传变异增大。本研究中, 用于研究的52份老芒麦来自5个不同的地理区域, 且它们具有不同的生态地理条件。有研究认为, 引物平均多态信息含量值(PIC)与群体遗传变异程度正相关[39]。本研究所用20对EST-SSR引物PIC值范围为0.260.50, 平均值为0.43, 表明供试老芒麦材料间遗传变异程度较高。同时, 这些引物扩增出204条带, 其中176条具多态性。以上结果及前人的研究分析均表明EST-SSR标记用于评价老芒麦遗传多样性是一种有效的工具。
关于野生材料与品种/栽培材料间遗传多样性比较的研究未见报道。本研究结果表明, 与品种/栽培材料(PPB=50.98%, I=0.234 7, H=0.151 0, Na=1.509 8)相比, 野生材料(PPB=85.29%, I=0.296 6, H=0.179 8, Na=1.852 9)具有更高的遗传多样性。有研究表明, 与野生材料相比, 品种/栽培材料的遗传多样性较低, 如荔枝(Litchi chinensis)[40]、水稻(Oryza rufipogon)[41]、大豆[42]和珍珠粟(Pennisetum glaucum)[43]等。研究者曾指出与目的农艺性状相关的大多数位点(如生育期、叶面积和产量等)在育种进程中被选择并保留下来, 但也有很多基因在此过程中丢失[44]。
单株选择法或系谱法, 是自花授粉植物培育新品种的一种基本方法[45]。前人报道指出, 单株选择法可以从混合系中分离出纯系, 并可以在其子代保留最好的基因型[46]。因此, 作为一种自花授粉植物, 老芒麦在育种进程中可能因多次单株选择, 导致杂合子的数目减少而纯合子增加[47]。以上因素降低了老芒麦品种/栽培材料的遗传多样性。相比之下, 野生种质材料, 因此它们具有更高的遗传多样性[48, 49]。因此, 具有丰富遗传多样性的野生材料在今后的育种工作中应作为重要的遗传资源。
本研究采用EST-SSR标记构建老芒麦指纹图谱, 能够区别不同品种及栽培材料的遗传特异性, 同时也为国内老芒麦品种知识产权和育种家的权益提供保障。
其他的研究已证明国内的老芒麦种质资源丰富且具多样性, 可以通过不同的标记方法揭示其丰富的表型性状和较高的遗传多样性[2, 3, 4, 5, 6, 18, 19]。本研究发现, 与品种/栽培材料比较, 野生材料具有更高的遗传多样性, 且地理类群内具有更大的遗传变异。研究表明, 近年来生态环境日益恶化加之自然灾害频发, 造成大量野生老芒麦种质资源急剧下降[50]。同时, 落后、单一的育种方法及种质收集保存不充分, 使得国内只有少数的老芒麦品种[51, 52]。因此, 需要更加关注原位保护和非原位保护, 以及特殊地理类群的老芒麦种质的收集与保护, 从而为优异种质筛选和育种奠定基础。
综上, EST-SSR分子标记是研究老芒麦种质遗传多样性的有效方法, 由此也证明我国老芒麦种质具有丰富的遗传变异, 这在野生老芒麦材料中体现的尤为明显。为此, 在开展老芒麦育种研究工作的同时, 也更应注重老芒麦种质资源的收集、保护和利用等。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
利用EST-SSR标记构建中国老芒麦品种DNA指纹图谱及种质遗传多样性
张俊超, 谢文刚, 赵旭红, 张宗瑜, 赵永强, 王彦荣
EST-SSR marker based DNA fingerprinting for cultivar identification and genetic diversity analysis of Elymus sibiricus germplasm in China
Zhang Jun-chao, Xie Wen-gang, Zhao Xu-hong, Zhang Zong-yu, Zhao Yong-qiang, Wang Yan-rong