
{kind=link}
{kind=link}
{kind=link}
青藏高原东北缘高寒草甸珠芽蓼斑块植被构成和种间关联
[魏斌 , 王莹, 关士琪, 高红娟, 孔杨云, 于应文]
, 王莹, 关士琪, 高红娟, 孔杨云, 于应文]
, 王莹, 关士琪, 高红娟, 孔杨云, 于应文]
|
|


第一作者:魏斌(1996-),男,甘肃永登人,在读本科生。E-mail:weib14@lzu.edu.cn
群落植物种构成和种间关系反映植物种群和群落分布格局上的互作关系。通过对珠芽蓼( Polygonum viviparum)斑块及其对照区域群落植物种重要值、物种多样性指数、生物量构成及植物种种间关联Jaccard指数和Spearman秩相关进行分析,揭示高寒草甸演替过程中的生态学机制。结果表明,珠芽蓼斑块的样方物种数显著高于对照( P<0.05),二者类群生物量与其重要值构成变化规律类似。珠芽蓼斑块的植被构成以杂类草和禾草为主,其占绿色生物量的60.9%和22.1%,且珠芽蓼种群约占绿色生物量的1/3;而对照则以禾草和杂类草为主,分别占绿色生物量的53.3%和27.1%。珠芽蓼斑块及对照群落中,显著关联的种对数分别占各自总种对数的16.4%和33.0%,多数植物种对的种间联结较松散,种对间独立性较强,二者正相关与负相关种对数比分别为0.83和1.40。珠芽蓼斑块群落处于稳定阶段,珠芽蓼斑块的形成,能增强高寒草甸的群落稳定性。
The vegetation composition and interspecific relationship reflect the interaction relationship of spatial distribution pattern among plant populations and communities. We analyzed the important value, diversity index of species, biomass composition, the Jaccard association index and spearman rank correlation coefficient of main species growing on patch in Polygonum viviparum alpine meadow ( P. viviparum patch) and control (CK), in order to reveal the ecological mechanism of succession process in Alpine meadow. The results indicated that number of species in P. viviparum patch was bigeer than that in CK, and the change pattern of each group was similar between important value and biomass composition. P. viviparum patch dominated mainly by forbs and grasses, accounting for 60.9% and 22.1% of the green biomass, respectively, and P. viviparum accounted for about one third of the green biomass; while it dominated mainly by grasses and forbs, accounting for 53.3% and 27.1% of the green biomass, respectively. In P. viviparum patch and CK, the significantly associated species-pairs accounted for 16.4% and 33.0% of their respective total species-pairs. The interspecific association of most species-pairs was not significant, and the independence between species-pairs was stronger. The ratios of negative and positive correlations were 0.83 and 1.40, respectively. Plant communities of P. viviparum patch was a stable stage. The formation of P. viviparum patch can strengthen community stable of alpine meadow.
高寒草甸是青藏高原主要草地类型之一, 它不仅是支撑高原畜牧业发展、维系地方经济发展的物质基础, 而且对长江中下游平原和黄河、长江流域地下水、地表水的丰歉以及防止风、旱、涝、沙尘暴、水土流失等自然灾害的发生, 起着不可替代的屏障作用[1]。然而, 由于受气候、人口和经营方式等的影响, 高寒草地的退化已威胁到当地生态环境维持、生物多样性保护和畜牧业经济发展。因此, 退化高寒草甸的恢复治理是管理青藏高原高寒生态系统的关键。草地植被斑块状分布是天然放牧草地的基本特征, 是植被对放牧和气候因素长期适应的结果[2, 3]。植物种间联结是群落的本质特征之一, 是指不同植物种在空间分布上的相关性, 因群落生境差异影响物种分布而引起[4, 5]。研究草地群落中斑块的群落构成特征和种间关联, 反映植物物种空间上的相互关系, 对于高寒草甸植被演替动态和稳定性机制的揭示具有重要意义。
近年来, 在青藏高原高寒草甸上, 诸多学者针对嵩草(Kobresia)和禾草斑块的植被特征[2, 6], 双子叶斑块的植被与土壤养分特征关系[7], 禾草、莎草和双子叶植被斑块的植被构成及植物种空间分布[3, 8], 粪斑和鼠丘斑块植被和土壤养分变化[9, 10, 11]等进行了研究, 这为草地植被斑块的深入研究提供了理论基础和分析方法。珠芽蓼(Polygonum viviparum)作为青藏高原高寒草甸草地常见植物种之一, 常以斑块形式存在于草地中, 在高寒草甸演替中起重要作用。诸多学者对珠芽蓼的药用价值和化学成分[12]、生理生化特性[13]以及繁殖特性[14]等方面进行了系统研究, 但就珠芽蓼斑块植物种间关系的研究相对缺乏, 虽然顾宁等[15]对山西芦芽山珠芽蓼群落中主要植物种的种间关联进行了分析, 但尚无放牧条件下珠芽蓼斑块的植物种间关系研究。本研究以青藏高原东北缘高寒草甸草地中珠芽蓼斑块为研究对象, 采用Jaccard种间关联度指数及Spearman秩相关方法, 分析珠芽蓼斑块及其对照区域主要植物种的构成和种间关联, 旨为珠芽蓼斑块的演替动态及稳定性机制的深入揭示提供理论基础和实践依据。
研究区位于青藏高原东北缘的甘肃省天祝藏族自治县抓喜秀龙乡的高寒草甸上, 地理坐标为102° 40'-102° 47' E, 37° 11'-37° 14' N, 海拔2 960 m。年均气温-0.1 ℃, 1月均温-18.3 ℃, 7月均温12.7 ℃, > 0 ℃年积温1 380 ℃· d。年降水量416 mm, 无绝对无霜期, 仅分冷热两季, 土壤为亚高山草甸土。草地主要植物种有矮嵩草(Kobresia humilis)、线叶嵩草(K. capillifolia)、珠芽蓼、西北针茅(Stipa krylovii)、阴山扁蓿豆(Melilotoids ruthenica var. inschanica)和球花蒿(Artemisia smithii)等[9]。
于2015年7月中旬, 在多年放牧牦牛和藏羊、草地利用率为80%~90%的冷季牧场(放牧期一般为11月-翌年4月末)的高寒草甸上, 设置面积为0.3~0.5 hm2的样地3块。
1.3.1 群落构成特征 2015年7月下旬-8月下旬, 在各样地内, 随机选择珠芽蓼斑块17个, 珠芽蓼斑块的确定依据为在借鉴牟晓明等[3]和张卫国等[10]的草地斑块划分依据及高寒草甸珠芽蓼斑块实际调查基础上, 将面积在1.0 m2以上的与周围植物群落组成明显不同、珠芽蓼种群聚集的斑块状草地命名为珠芽蓼斑块。以珠芽蓼斑块为中心, 用0.5 m× 0.5 m的样方, 调查测定各珠芽蓼斑块和配对的距离珠芽蓼斑块边界约1.0 m处对照区域内所有植物种的高度、盖度和地上生物量。先将收获的地上生物量(total biomass, TB)按绿色(活)物质(green biomass, GB)和死物质(dead biomass, DB)分开, 再将绿色物质按不同植物种分开, 然后置入烘箱烘干称重。按公式IV=(RC+RH+RB)/3计算植物种重要值(important value), 式中:IV 为植物种重要值, RC、RH和RB分别为植物种的相对盖度(样方内某种的盖度/样方内所有种的盖度之和)、相对高度(样方内某种的高度/样方内所有种的高度之和)和相对生物量(样方内某种的生物量/样方内所有种的生物量之和)。
利用样方调查数据, 按公式[4]DGI=S/lnA计算物种α 多样性指数DGI(Gleason指数), 式中:S为群落中的物种数目, A为样方面积; 按公式[4]β w=(S/ma)-1计算物种β 多样性指数β w(Whittaker指数), 式中:S为研究系统中观测物种总数, ma为各样方或样本的平均物种数。
1.3.2 植物种间关联度Jaccard指数 Jaccard指数反映的是物种共同出现的百分率。采用Jaccard指数[5](简称JI指数)测定物种间关联程度, 以2× 2列关联表为基础计算, 其公式为JI=a/(a+b+c)。式中:a为物种A和物种B同时出现的样方数; b和c分别为物种A或物种B出现一种的样方数。当a=0时, 表示种A和种B不同时出现在同一样方内, 即种A和种B完全独立或相异; 当a=S(样方数)时, b=c=0, 取值为1, 表示物种A、B同时出现在所有样方中, 即种A与种B绝对关联; 当b=0时, 表示种B与种A完全关联, 即种A随种B的出现而出现, 而种B可以单独出现; 当c=0时, 表示种A与种B完全关联, 即种B随种A的出现而出现, 而种A可以单独出现。本研究在借鉴张金屯[5]的划分方法基础上, 将JI指数划分为4个等级, 对珠芽蓼斑块主要植物种种间关联进行客观分析, 0.75< JI≤ 1为强联结, 0.5< JI≤ 0.75为次强联结, 0.25< JI≤ 0.5为弱联结, 0≤ JI≤ 0.25为无联结。
1.3.3 植物种间Spearman秩相关分析 Spearman秩相关系数[5]属于非参数性检验, 由植物种重要值计算。其公式为:
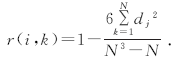
式中:r(i, k)为物种i与物种k的相关系数, N为样方数, dj=(xij-xkj), xij、xkj分别为物种i和物种k在样方j中的秩。
用Excel进行数据处理及制图。SPSS 16.0软件的One-Way ANOVA对斑块和对照区域的植物群落特征和生物量进行方差分析(F-检验)和显著性检验, 数据用均值± 标准误差表示。
群落主要植物种重要值显示, 珠芽蓼斑块与对照区域草地植物种构成差异很大(表1)。珠芽蓼斑块以珠芽蓼为优势种, 其重要值为0.351; 球花蒿为亚优势种, 其重要值为0.168, 优势种约为亚优势种的两倍; 垂穗披碱草、线叶嵩草、矮嵩草、西北针茅、秦艽、早熟禾和赖草等为主要伴生种; 对照的优势种为西北针茅和矮嵩草, 其重要值分别为0.179和0.152, 对照无明显亚优势种, 早熟禾、球花蒿、垂穗披碱草、艹洽草、秦艽、赖草和兰花韭等为主要伴生种。
| 表1 珠芽蓼斑块群落主要植物种重要值 Table 1 The important value of P. viviparum patch and control (IV≥ 0.01) |
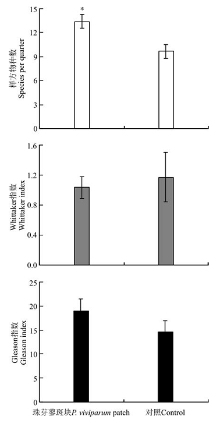
珠芽蓼斑块和对照草地的草层高度分别为14.83± 1.66和17.56± 3.60 cm, 植被盖度分别为98.6%± 1.15%和94.8%± 3.58%, 且二者均在珠芽蓼斑块和对照区域之间差异不显著(P> 0.05)。珠芽蓼斑块的群落物种数显著高于对照区域(P< 0.05), 其Gleason指数和Whittaker指数在珠芽蓼珠芽蓼斑块和对照区域之间差异不显著(P> 0.05)(图1)。说明, 珠芽蓼斑块有利于物种多样性的形成。
 | 图1 珠芽蓼斑块和对照区域植物物种多样性注:* , P< 0.05。Fig.1 Species diversity of P. viviparum patch and control |
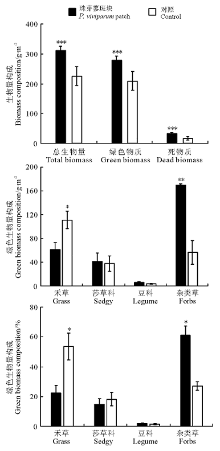
珠芽蓼斑块群落地上总生物量、绿色物质量和死物质量均极显著高于对照区域的(P< 0.001), 其禾草生物量及其比例表现为珠芽蓼斑块显著小于对照(P< 0.05), 莎草和豆科植物生物量及其比例在珠芽蓼斑块和对照之间均无显著差异(P> 0.05)。珠芽蓼斑块群落地上绿色生物量主要由杂类草和禾草植物构成, 分别占绿色生物量的60.9%和22.1%, 且珠芽蓼种群几乎占绿色生物量的1/3; 而对照区域以禾草和杂类草植物为主, 分别占绿色生物量的53.3%和27.1%。珠芽蓼斑块和对照区域草地类群生物量比例与其重要值构成变化规律类似。
 | 图2 珠芽蓼斑块和对照区域的群落生物量构成注:* , P< 0.05; * * , P< 0.01; * * * , P< 0.001。Fig.2 Biomass composition of P. viviparum patch and control |
珠芽蓼斑块及其对照区域主要植物种种间Jaccard指数显示, 珠芽蓼斑块中强联结种对数为21对, 次强联结种对数为30对, 弱联结种对数为4对, 无联结种对数为0对, 分别占总种对数的38.2%、54.5%、7.3%和0。对照中强联结种对数为9对, 次强联结种对数为36对, 弱联结种对数为23对, 无联结种对数为23对, 分别占总种数的9.9%、39.5%、25.3%和25.3%(表2)。说明, 珠芽蓼斑块植物种的种间关联性强, 群落处于稳定阶段。
| 表2 珠芽蓼斑块主要植物种种间Jaccard指数(JI) Table 2 The interspecific Jaccard indices of P. viviparum patch and control |
Spearman秩相关分析显示, 珠芽蓼斑块中, 正相关的种对数为25对, 其中极显著(P< 0.01)和显著(P< 0.05)相关的各两对, 不显著(P> 0.05)相关的21对; 负相关的种对数为 30对, 其中极显著相关的种两对, 显著相关的3对, 不显著相关的25对。对照区域, 正相关的种对数53对, 其中极显著相关的6对, 显著相关的11对, 不显著相关的36对; 负相关的种对数38对, 其中极显著相关的3对, 显著相关的10对, 不显著相关的25对。珠芽蓼斑块和对照区域中, 显著关联的种对数分别占各自总种对数的16.4%和33.0%, 其中显著、极显著正相关的种对比例分别为7.3%和18.7%, 显著、极显著负相关的种对比例分别为9.1%和14.3%。珠芽蓼斑块的55个种对和对照区域的91个种对中, 绝大多数种对的联结关系未达到显著水平, 种间联结较为松散, 种对间的独立性相对较强。珠芽蓼斑块中正联结种对占总种对数的45.45%, 负联结种对占总种对数的54.55%, 正负关联比为0.83; 而对照区域中正联结种对占总种对数的58.24%, 负联结种对占总种对数的41.76%, 正负关联比为1.40。说明, 珠芽蓼斑块中, 多数植物种对间联结性较弱, 种间独立性强, 两个对照的植物种对间联结性较强, 两个群落处于稳定阶段, 这与主要植物种种间关联度Jaccard指数所得结果类似。
 | 图3 珠芽蓼斑块和对照区域主要植物种Spearman秩相关分析半矩阵图Fig.3 Semi-matrix of spearman rank correlation analysis of P. viviparum patch and control |
本研究珠芽蓼斑块主要植物种种间关联度Jaccard指数和Spearman秩相关整体结果均显示珠芽蓼斑块群落处于稳定阶段。一方面由于珠芽蓼斑块具绝对优势种, 其植物种优势度构成模式更利于群落的稳定[16]; 另一方面由于珠芽蓼斑块中的珠芽蓼因富含单宁等物质, 家畜不喜采食[12], 牛羊仅在夏末秋初采食其花果[17], 且珠芽蓼斑块中可食植物如禾草等比对照区域的低, 从而放牧家畜对珠芽蓼斑块的整体采食程度低, 利于其群落稳定性的维持。而对照区域优势种西北针茅和矮嵩草的优势度较低, 无明显亚优势种, 且因家畜采食程度高而更易发生演替。这进一步表明, 珠芽蓼斑块中植物种对之间的竞争整体相对较弱, 群落成熟程度较高, 各物种间以相互独立的关系求得资源的充分利用, 从而保持斑块物种间的稳定发展; 而对照群落中植物种间的整体竞争相对激烈, 群落处于演替初、中期阶段, 群落容易发生演替变化。本研究珠芽蓼斑块群落的物种数和Jaccard关联度均高于对照区域, 可能是因为珠芽蓼斑块内群落稳定性较强, 群落内种间竞争比对照区域小, 而利于物种多样性的形成。由此说明, 家畜不喜食或采食程度低的植被斑块的形成更利于群落稳定性和物种多样性的形成。
植物种间正关联, 主要是由于它们的生物学特性相近、对生境具有相似的生态适应性[18, 19]。本研究的珠芽蓼斑块中, 亚优势种球花蒿与赖草种对间呈显著正相关, 它们均具耐旱、耐寒、耐贫瘠, 生态适应性较强的特征, 从而二者具有相似的生境和利用资源的特征; 对照区域中优势种西北针茅、矮嵩草与其它植物无显著正相关关系。植物种间负关联, 主要是由于生物学特性不同, 对生境具有不同的生态适应性或相互竞争所致[20]。本研究珠芽蓼斑块中, 优势种珠芽蓼与伴生种线叶嵩草、矮嵩草、甘肃棘豆之间的关联均呈显著负相关, 是由于这些伴生种根系细弱入土较浅, 对土壤水分和养分的吸收能力较弱, 而珠芽蓼具粗壮肥厚根茎[14], 对水分和养分吸收的竞争能力较强, 且其总酚类和黄酮含量高, 抗氧化能力强[21], 从而珠芽蓼对环境的适应性更强, 由此, 珠芽蓼与这些植物种之间存在明显排斥性; 对照区域的优势种西北针茅、矮嵩草与早熟禾、秦艽、兰花韭、喉毛花、唐松草种对间呈显著或极显著负相关, 是由于西北针茅和矮嵩草与这些伴生种之间具有不同生境适应性和资源利用特征, 相互之间的竞争或生态适应性不同, 即呈现相互排斥。此外, 本研究珠芽蓼斑块和对照区域正相关与负相关的植物种对数均存在, 这与顾宁等[15]在山西芦芽山珠芽蓼群落植物种间关联整体呈正关联的研究结果存在分异, 可能与其珠芽蓼群落无放牧利用有关。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|