




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
高寒草原群落特征对甘肃马鹿冬季放牧的响应
[胡俊奇, 陈先江, 侯扶江 ]
]
]
|
|


第一作者:胡俊奇(1989-),男,河南许昌人,在读硕士生,主要从事草地家畜互作研究。E-mail:hjq891204@126.com
在肃南裕固族自治县甘肃马鹿( Cervus elaphus kansuensis)冬季牧场,采用巢式样方法调查草地植被群落特征随放牧压力变化的响应,揭示马鹿在放牧系统中的地位和功能,对保护、恢复与重建退化放牧草地生态系统具有重要作用。结果表明,株高<10 cm物种的密度、盖度、地上现存量占总密度、总盖度、地上总现存量的比例均随放牧率增大而增大,并与放牧率呈线性相关。凋落物枯黄期(10月中旬)最多,凋落物量与放牧率呈极显著负相关( P<0.01)。放牧促进植物个体小型化,一定放牧率内,物种丰富度随放牧强度增大而降低,且与植物个体小型化有复杂的互作关系。
The nest-style method was used to investigate the response of plant community characteristics to grazing pressure to analyze the position and function of wapiti in the grazing system in the winter pasture of Gansu wapiti in Sunan Yugu National Autonomous County where wapiti has an important effect on protecting, restoring and reconstructing of degenerated grassland ecosystem. This study showed that the plant density, coverage and aboveground biomass increased with the increase of grazing intensity, and they were a linear positive correlation with grazing rate when plant height was below 10 cm. The plant litter biomass reached a peak in wither period (middle October), and it had a significant negative correlation with the grazing rate. The grazing made the plant miniaturization, and the species richness had a decrease trend with the increasing grazing intensity under given grazing rate. A complex interaction relationship between grazing intensity and plant miniaturization was observed in this study.
放牧是草原最重要的利用方式[1, 2], 也是影响草原群落结构与功能的主要途径[3]。放牧对于草原生物多样性和牧草个体形态的作用, 始终是研究的热点之一[4]。 放牧主要通过家畜的采食和践踏对牧草个体形态产生影响。家畜采食导致牧草冠幅直径变小, 对空间和阳光的利用程度降低, 生长变慢, 个体趋于矮小; 家畜对牧草的践踏, 导致土壤理化性质发生变化, 在土壤表皮形成一层紧实层, 牧草根系分布变浅, 牧草对土壤水分和矿质养分的利用程度降低, 影响牧草生长, 植物个体趋于小型化。轻度和中度放牧增加群落物种丰富度, 促进植物个体小型化[5]。也有研究认为, 长期过度放牧引起草原群落退化, 降低物种丰富度, 导致个体植株变矮, 群落植物个体表现出小型化趋势[6]。但是, 牧草个体小型化与放牧率和生物多样性的定量关系尚缺乏必要的研究。
植物个体形态和物种多样性是反映群落特征的重要指标。祁连山高山草原因海拔高, 环境恶劣, 物种多样性对放牧的响应尤其敏感[7]。因此, 本研究以祁连山中段肃南马鹿冬季牧场为研究对象, 以不同放牧地的群落密度、盖度、地上现存量、物种丰富度为指标, 探讨祁连山高山草原群落特征对放牧的响应。
研究区位于甘肃省肃南裕固族自治县甘肃马鹿(Cervus elaphus kansuensis)养殖场, 地处祁连山中段北坡, 也是青藏高原的北坡东段, 是青藏高原畜牧区与河西走廊农耕区之间过渡的陡梯地带。地理坐标38.8° N, 99.6° E, 海拔2 850 m。年均温3.6 ℃, 年均降水量253.0 mm, 主要集中在6―9月, 年均蒸发量1 784.6 mm。土壤为山地栗钙土。牧草一般4月下旬返青, 7月上旬进入生长旺盛期, 9月中上旬开始枯黄。草地群落主要物种是紫花针茅(Stipa purpurea)、波伐早熟禾(Poa poiphagorum)、扁穗冰草(Agropyron criststum)、矮嵩草(Kobresia humilis)、甘肃苔草(Carex kansuensis)、短花针茅(S. breviflora)等, 退化地段主要有银灰旋花(Convolvulus ammannii)和醉马草(Achnatherum inebrians)[8]。
根据草原综合顺序分类法, 草地类型属于寒温微干山地草原类[9]。草地农业系统类型主要是天然草地— 家畜生产系统。
甘肃马鹿有春秋季、夏季和冬季3个季节牧场, 研究区位于冬季牧场。冬季牧场地形开阔平坦, 整体坡度< 3° , 面积约160 hm2, 每年11月下旬-翌年4月放牧甘肃马鹿[9]。通过实地调查和询问牧民, 确认马鹿放牧采食路线, 同时结合草原植被状况, 以牧场进出口为起点, 向外呈放射状形成由高到低的牧压梯度。分别在距牧场出入口0、300、600、900、1 200、1 500 m处设6个样地, 每个样地取样面积约1 hm2, 标明边界。
用望远镜远距离观测马鹿在样地的出现次数; 从出牧到归牧(北京时间08:30-16:30)每2 h观测1次, 每天观测4次, 每次每样地观测不少于15 min, 连续观测15 d。计算家畜一天中在样地出现的总频次; 一个放牧季观测3次以上。
6个样地放牧率(stocking rate, SR)根据王化等[9]、Zhang等[10]和Yuan和Hou[11]的公式计算:
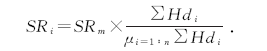
式中:SRi为样地i的放牧率; SRm为整个牧场的平均放牧率, 一般为家畜单位与牧场面积的比值, 本研究中再乘以放牧时间; ∑ Hdi为观测期间样地i出现的家畜频率次数总和; μ i=1∶ n∑ Hdi为观测期间所有样地出现的家畜次数总和的平均值; n为样地总数。测定各年龄段马鹿的活体重, 换算为家畜单位。本研究中, 一个家畜单位(animal unit, AU)为一头成年甘肃马鹿母鹿(活体重200 kg左右), 带一只哺乳期的小鹿。根据体量或采食量, 将各年龄阶段马鹿换算为家畜单位。从牧场出入口向远处的6个样地对应的放牧率AUM(animal unit× month, 放牧家畜单位数与放牧月份的乘积)分别为6.90、4.85、3.45、2.45、1.45和1.00 。
1999年采用限定随机取样法[8]在每个样地做10个0.5 m× 0.5 m的样方, 测定每种植物的现存量, 统计各样方中的植物种数和每种植物的株丛密度。
冬季牧场群落在营养生长期明显分为两个亚层, 即> 10 cm亚层和< 10 cm亚层, 前者主要是紫花针茅、赖草(Leymus secalinus)、醉马草、扁穗冰草、波伐早熟禾等禾草, 后者主要是银灰旋花、冷蒿(Artemisia frigida)和二裂委陵菜(Potentilla bifurca)等双子叶类。在每样地, 每种植物随机测定30个个体的株高, 根据种群平均高度分为< 10 cm物种和> 10 cm物种。
分别在5月上旬、6月中旬、7月下旬、9月上旬、10月中旬观测5次。
试验数据用Excel进行初步整理, 用SPSS 20.0进行统计分析。
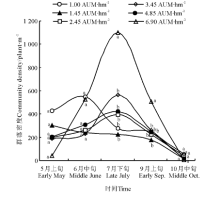
试验期间, 6.90、4.85、3.45、2.45 AUM· hm-2样地草原群落密度的季节动态基本呈单峰曲线(图1), 均以7月下旬最高。7月下旬和9月上旬, 放牧率较大(6.90)的样地群落密度显著高于其他样地(P< 0.05), 群落密度与放牧率呈显著正相关(表1), 这是由于银灰旋花、黑褐苔草(C. atrofusca)等小个体物种的密度大, 占绝对优势所导致。5月上旬, 放牧率较小的1.00和1.45 AUM· hm-2样地的群落密度较高, 且放牧率与群落密度呈负相关(表1), 说明放牧率大对翌年牧草返青不利。
 | 图1 冬季牧场不同放牧率下的植物群落密度月动态Fig.1 Monthly dynamics of community density in the winter pasture under different stocking rate 注:不同字母表示同一时间不同放牧率之间差异显著(P< 0.05)。 Note: Different lower case letters at the same month indicate significant difference among different stocking rates at 0.05 level. |
| 表1 冬季牧场群落密度(Y)与放牧率(X)的关系 Table 1 Relationship between community density (Y) and stocking rate (X) in the winter pasture |
生长旺季, 群落下层(平均株高< 10 cm)物种的种群密度与放牧率呈极显著正相关(图2)。株高< 10 cm物种的密度占总密度的比例与放牧强度呈极显著正相关(图3), 放牧率每增加1个单位, 比例上升7.4%左右; 群落下层(平均株高< 10 cm)物种的盖度占群落总盖度的比例也随放牧强度增大而上升。表明, 放牧较重利于高寒草原小个体物种的形成, 即促进种群个体小型化。
 | 图2 7月下旬株高< 10 cm物种的密度和盖度与放牧率的关系Fig.2 Relationship between the density, coverage of species of plant height < 10 cm and stocking rate in late July |
 | 图3 7月下旬株高< 10 cm物种的密度、盖度及地上现存量比例与放牧率的关系Fig.3 Relationship between density, coverage, standing biomass percentage of species of plant height < 10 cm and stocking rate in late July |
群落盖度对群落冠层微气候形成有重要作用, 直接影响群落生长[13]。本研究中群落盖度为各种群盖度之和, 即分盖度之和。
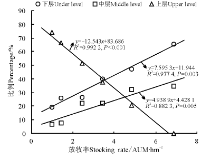
7月下旬群落层次可以细分为0―10 cm(底层, under level)、10-20 cm(中层, middle level)和≥ 20 cm(上层, upper level)3个亚层。随放牧强度增大, 群落下层和中层物种的盖度占总盖度的比例呈增加趋势, 而上层物种盖度占总盖度的比例变化趋势相反(图4), 表明放牧强度越大, 越有利于中小个体物种的种群, 即促进种群个体的小型化。
 | 图4 冬季牧场群落冠层结构与放牧率的关系Fig.4 Relationship between community canopy structure and stocking rate in winter pasture |
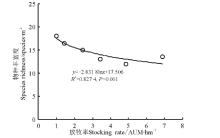
放牧率≤ 4.85 AUM· hm-2, 物种丰富度随着放牧强度增大呈现递减趋势; 放牧率最大的6.90 AUM· hm-2样地, 物种丰富度反而高于放牧较轻的4.85和3.45 AUM· hm-2样地(图5), 原因是放牧较重样地家畜活动集中、排泄物较多, 家畜从牧场其它区域携带较多种子、特别是一年生种子到此, 可以利用较为丰富的排泄物养分进行生长繁殖。物种丰富度与放牧率的关系可以用抛物线拟合(P=0.001), 理论上放牧率为5.04 AUM· hm-2时, 物种丰富度最低, 为12.17种· m-2(图5)。
 | 图5 物种丰富度与放牧率的关系Fig.5 Relationship between species richness and stocking rate |
物种丰富度随群落小型化各项指标(平均株高< 10 cm物种的种群密度、盖度及其占总密度、总盖度的比例)变化呈先下降后上升趋势(图6)。说明放牧率对于物种丰富度和群落物种小型化的作用机制有相似之处, 物种丰富度与小型化之间具有复杂的互作关系。
 | 图6 物种丰富度与草层高度< 10 cm物种的密度和盖度的关系Fig.6 Relationship between species richness and density, coverage of plant species with height < 10 cm |
本研究表明, 甘肃马鹿冬季放牧促进祁连山高寒草原植物个体小型化, 这与近年国内外的研究结论类似(图3)。青海高寒草原在40年间, 紫花针茅株高下降了5.89~11 cm, 降幅为42.3%~44.5%, 个体形态表现出小型化趋势[12]; 一般认为, 退化的草原群落, 特别是丛生的植物种群, 能够通过小型化抵御家畜的采食[13, 14, 15]。
研究植物个体小型化对于揭示草原群落退化机制有重要意义。植物地上部的小型化意味着个体对微气候资源(光、热、气等)的竞争能力降低[16], 不同植物种群小型化的差异导致草原空间和其它资源在植物种群间再分配, 这是植物种群在群落中优势地位消长的基础之一[17]。放牧促进植物个体小型化, 小个体种群会得到更多的资源和空间, 促进生长和繁殖, 种群数量增加, 优势度增大[18, 19]。
本研究群落物种数随放牧强度增大呈递减趋势, 且随放牧率增大, 降幅减小, 说明改进放牧管理能在一定范围内增加物种多样性, 与“ 中度干扰假说” 相一致[20]。但是, 在内蒙古荒漠草原的研究发现, 随着放牧强度的增加, 群落物种丰富度显著降低, 可能与荒漠草原植被种类少, 抗干扰能力较差有关[21]; 还有研究发现, 虽然放牧能增加物种多样性, 但也提高牧草中的木质素含量, 降低牧草品质[22]; 而且牛的放牧比羊更能增加草地物种多样性[23]。国外有研究表明, 草地物种多样性和土壤质地不仅与放牧有关, 也与放牧前土地利用方式有关[24]; 而且适度放牧可以增加植物多样性, 从而修复退化草地[25, 26]。
放牧对草原地上生物量的影响同样复杂[27]。国内外研究表明, 适度放牧可以显著提高巴塔哥尼亚干旱草原和黄土高原草原地上生物量[28, 29]; 过度放牧增加阿根廷恩佩德拉多草原地上生物量中绿色组织的比例(30%), 但地上总生物量减少[30]。本研究凋落物量随着放牧强度增大而减少, 与多数放牧研究结果相似[28, 29, 30, 31], 说明放牧较重不利于凋落物积累, 原因可能是放牧加速凋落物分解, 或降低凋落物的行程, 或二者兼有, 其机理尚需进一步研究。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|