
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
外源甜菜碱对盐胁迫下黑果枸杞种子萌发和幼苗保护酶活性的影响
[李善家1  , 韩多红
, 韩多红2 , 王恩军2 , 武燕1 ]
, 韩多红, 王恩军|
|


第一作者:李善家(1980-),男,甘肃兰州人,讲师,博士,主要从事干旱环境植物生理生态学研究。E-mail:lishanjia@lut.cn
为了探究外源甜菜碱对盐胁迫下黑果枸杞( Lycium ruthenium)种子萌发及抗氧化物酶系活性的影响,对采自于河西走廊黑河中游的黑果枸杞进行种子萌发及幼苗生长试验,测定其发芽率(GP)、发芽势(GV)、发芽指数(GI)、活力指数(VI)、相对盐害率和抗氧化物酶系[超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)]等指标。结果显示,25 mmol·L-1的NaCl处理,对黑果枸杞种子的萌发具有促进作用,50~300 mmol·L-1的NaCl处理则有抑制作用;黑果枸杞种子萌发的NaCl浓度的临界值是50 mmol·L-1,极限值是300 mmol·L-1;随着NaCl浓度的增加,SOD、POD和CAT活性均不同程度地表现为“先上升,后下降”的趋势。在添加外源甜菜碱处理后,各NaCl浓度处理下的SOD、POD和CAT活性均有不同程度的增加。综上表明,甜菜碱能有效减缓盐胁迫对黑果枸杞种子萌发及幼苗生长产生的伤害,提高种子及幼苗的抗盐能力。
In order to investigate the effects of exogenous betaine on seed germination, activities of antioxidase enzymes of Lycium ruthenium under NaCl stress. The L. ruthenium seed germination and seedling growth were studied in middle reaches of Heihe River of Hexi Corridor. Several physiological indexes, such as the germination percentage (GP), germination vigor (GV), germination index (GI), vigor index (VI), and relative salt damage rate, antioxidant enzymes (SOD, POD and CAT) were measured. The results indicated that, low NaCl concentration (25 mmol·L-1) could promote the seed germination but high NaCl concentration (50~300 mmol·L-1) could inhibit the seed germination. The NaCl concentration threshold for seed germination in L. ruthenium was 50 mmol·L-1, the maximum was 300 mmol·L-1; The SOD, POD and CAT activity showed a trend of first rising and then falling along with the salt concentration increasing. After the treatment by exogenous betaine, SOD, POD and CAT activity increased to some extent, but they had less decrease range under NaCl stress. Exogenous betaine could significantly alleviate the damages to the seeds and seedlings of L. ruthenium under NaCl stress and promote the salt resistance of the seeds and seedlings.
盐碱地是地球陆地上分布较广泛的一种土地资源, 随着现代工业和灌溉农业的发展, 环境持续恶化, 盐碱地的面积也在逐年扩大, 已成为全球重要的后备土地资源。我国从滨海到内陆, 从低地到高原都分布着不同类型的盐碱土壤, 它们主要分布于西北、华北、东北和滨海地区, 总面积达到2 000万hm2, 约占总耕地面积的10%[1]。黑河流域地处西北干旱区, 属于典型的内流盆地, 气候干燥, 蒸发量是降水量的35~77倍, 恶劣的自然条件和人为的不合理灌溉导致了严重的土壤盐渍化。如何利用已经盐渍化的土地, 寻找适合其生长的植物成为解决土地资源困境的主要途径。在盐胁迫作用下, 植物生长受到抑制, 光合作用减弱, 代谢失调, 严重时出现植株萎蔫, 甚至死亡。甘氨酸甜菜碱是一种在较大生理pH值范围内呈电中性的季铵类化合物, 极易溶于水, 广泛分布在植物、动物和细菌体内。近年来, 外源甜菜碱与植物抗逆性的研究日益受到人们的重视, 在盐胁迫、水分及低温等胁迫下, 一些植物均检测到较高浓度的甜菜碱类化合物的积累[2]。甜菜碱作为渗透调节剂、酶的保护剂, 其积累使许多代谢关键酶在渗透胁迫下能继续保持活性, 在一定程度上保持了盐胁迫下细胞膜的完整性[3]。
黑果枸杞(Lycium ruthenium), 为茄科(Solanaceae)枸杞属多年生耐盐、抗旱植物, 多分布于我国西北地区的盐碱土荒地、盐化沙地、路旁等各种盐渍化土壤或荒漠环境。果实入药, 其味甘、性平, 是民族医药中的常用药材, 据《晶珠草本》、《四部医典》记载, 藏医以其成熟的果实入药, 治疗心热病、心脏病、月经不调、停经等病症[4]; 而《维吾尔药志》中将其果实和根皮治疗尿道结石、癣疥、齿龈出血等病症, 具有补肾益精、养肝明目、补血安神、生津止渴、润肺止咳、清热除蒸、清肺降火、补虚等功效[5]。其成熟浆果中富含紫红色素, 又属于珍稀的天然花色苷类色素资源, 具有清除自由基、抗氧化的功能[6]。目前, 对黑果枸杞的研究主要集中在天然花色苷色素和多糖提取工艺及药理研究方面[7, 8, 9], 部分学者也对其种子萌发的耐盐性进行了研究[10, 11, 12]。但对于盐胁迫下添加甜菜碱等方面的研究鲜有报道, 为此, 本研究通过分析盐胁迫下外源甜菜碱处理对黑果枸杞种子萌发和幼苗生长的影响, 探讨黑果枸杞在盐胁迫下的耐受性及其阈值, 旨为河西走廊大片盐碱化土地上的引种驯化、培育保护和大面积种植等提供理论依据。
黑果枸杞成熟种子于2013年9月采自甘肃省张掖市荒漠盐碱地, 为多年生野生种。将种子洗净后, 自然风干, 随即开始试验。
1.2.1 种子萌发指标的测定 选择籽粒饱满、大小一致的种子, 用5%的NaClO消毒3 min, 蒸馏水冲洗6次。设定NaCl的浓度为0、25、50、100、150、200、300 mmol· L-1共7个处理, 以NaCl浓度0为对照(CK), 添加的甜菜碱浓度为10 mmol· L-1(表1)。依照国际种子检验规程, 发芽床采用滤纸法, 将吸胀后的种子置于垫有两层滤纸的培养皿中, 分别加入配制好的处理液, 每个培养皿中放50粒种子, 每个处理均设3次重复, 置于人工气候箱中, 设定温度为25 ℃, 光照时间为10 h, 光照强度为2 000 lx。每天观察统计萌发数, 第3天计算发芽势, 第13天计算发芽率、发芽指数、活力指数和相对盐害率。
| 表1 盐胁迫及外源甜菜碱对黑果枸杞种子及幼苗的试验处理 Table 1 Experiment treatments on seed and seedings of Lycium ruthenium under NaCl and exogenous glycinebetaine |
发芽势(Germination vigor, GV)=(3 d内发芽种子数/供试所有种子数)× 100%。
发芽率(Germination percentage, GP)=(13 d内发芽种子数/供试所有种子数)× 100%。
发芽指数(Germination index, GI)=∑ (Gt / Dt)。;
活力指数(Vigor index, VI)=S× Gi。
式中, Gt为t天的发芽数, Dt为发芽日数, S为平均根长, 根长使用游标卡尺进行测量。
相对盐害率=(对照发芽率-处理发芽率)/对照发芽率× 100%。
1.2.2 幼苗抗氧化酶活性的测定 选取长势良好的幼苗, 将幼苗从培养皿中取出, 根部用无菌蒸馏水冲洗干净, 移入装有蛭石的育苗钵(直径15 cm× 高5 cm)内, 用Hoagland培养液进行浇灌, 在幼苗二叶期时定苗, 每钵留生长一致的幼苗15株, 3个重复, 50 d后进行处理。每天每钵分早晚加5 mL处理液, 以蒸馏水为对照。处理7 d后测定保护酶的活性。过氧化物酶(POD)采用愈创木酚法测量[13], 以每分钟吸光度的变化表示酶活力的大小, 即以每分钟A值减小0.01定义为1个酶活力单位(U); 超氧化物歧化酶(SOD)采用氮蓝四唑显色法(NBT)测定[13], 以抑制NBT光化还原50%所需的酶量为1个酶活力单位(U), 然后再计算出酶活力。过氧化氢酶(CAT)采用紫外吸收法测定[13], 以每分钟A值减小0.1定义为1个酶活力单位(U)。
采用SPSS 17.0对所测数据统计分析, 用平均值和标准误表示测定结果, 对数据进行单因素方差分析(P< 0.05), 并用Duncan法对各测定数据进行多重比较; 采用Excel 2010制图。
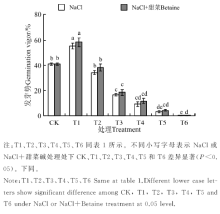
不同浓度的盐胁迫都对黑果枸杞的种子萌发产生了影响, 25 mmol· L-1处理有一定的促进作用, 50~300 mmol· L-1处理有抑制作用。在NaCl、NaCl+甜菜碱处理下(图1), T1的发芽势分别为55.11%、58.17%, 均显著高于CK(40.83%)(P< 0.05); T2与CK间无显著差异(P> 0.05)。T3~T6, 各处理的发芽率随处理浓度的增大而降低, 且显著低于CK(P< 0.05); 在T6处理中, 发芽势为0。NaCl+甜菜碱处理下, 各处理的发芽势与NaCl胁迫下相比没有明显的增大, 说明添加外源甜菜碱对种子发芽整齐度影响不大。
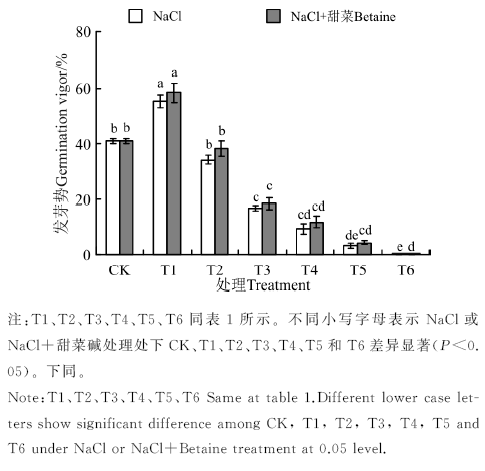 | 图1 不同处理下黑果枸杞种子的发芽率Fig.1 Germination rate of Lycium ruthenium seeds under different treatments |
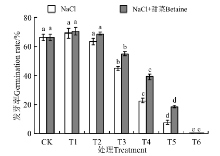
在NaCl处理下(图2), 25 mmol· L-1的发芽率(69.17%)高于CK(66.25%), 50 mmol· L-1(63.33%)的低于CK, 但25、50 mmol· L-1与CK间无显著差异(P> 0.05)。100~300 mmol· L-1的发芽率均显著低于CK(P< 0.05), 且随浓度的增大而降低, 在300 mmol· L-1时为0。在NaCl+甜菜碱处理下, 25 mmol· L-1(70.51%)、50 mmol· L-1(68.68%)的发芽率均高于CK(66.25%), 但与CK间无显著性差异(P> 0.05)。100~300 mmol· L-1, 各处理的发芽率也随处理浓度的增大而降低, 且显著低于CK(P< 0.05)。NaCl+甜菜碱处理下, 50~300 mmol· L-1的发芽率均比NaCl处理下的高, 且随着浓度的增大下降的趋势变缓。
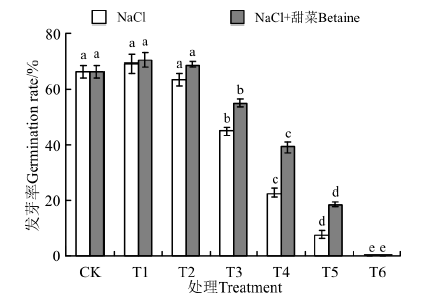 | 图2 不同处理下黑果枸杞种子的发芽势Fig.2 Germination vigor of Lycium ruthenium seeds under different treatments |
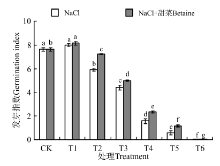
在NaCl处理下(图3), T1的发芽指数与CK间差异不显著(P> 0.05); T2~T6的发芽指数均显著低于CK(P< 0.05), 且随浓度的增大而降低, 在T6时为0。在NaCl+甜菜碱处理下, T1的发芽指数(8.13)显著高于CK(7.63), T2~T6的发芽指数均显著低于CK。
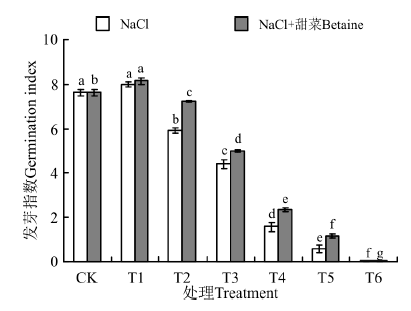 | 图3 不同处理下黑果枸杞种子的发芽指数Fig.3 Germination index of Lycium ruthenium seeds under different treatments |
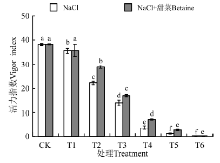
在NaCl、NaCl+甜菜碱处理下(图4), T1~T6的活力指数均低于CK, 并随浓度的增大而降低; 在NaCl处理下, 各处理与CK间存在显著性差异(P< 0.05)。但在NaCl+甜菜碱处理下, 各处理的活力指数与NaCl胁迫下相比均有不同程度的增大, 说明外源甜菜碱可缓解盐胁迫对种子萌发的影响。
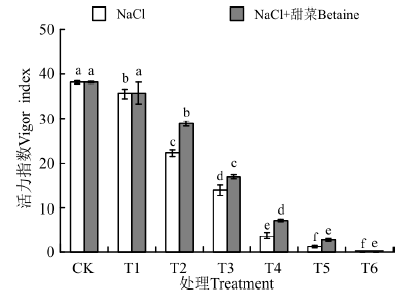 | 图4 不同处理下黑果枸杞种子的活力指数Fig.4 Vigor index of Lycium ruthenium seeds under different treatments |
在NaCl+甜菜碱处理下, T2~T6的发芽率均比NaCl处理下的高, 但各处理的发芽势与NaCl胁迫下相比没有明显的增大, 说明添加外源甜菜碱对种子发芽整齐度影响不大; 发芽率, 发芽势、发芽指数和活力指数均有所增大(图1-4)。
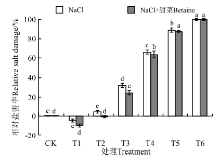
在NaCl处理下(图5), T1的相对盐害指数最小, 为-4.4%, T6最大, 为100%; T1、T2与CK之间没有显著性差异(P> 0.05), T3~T6均显著高于CK(P< 0.05)。在NaCl+甜菜碱处理下, T1的相对盐害指数更小, 为-9.47, T6最大, 为100%; T1、T2与CK之间没有显著性差异(P> 0.05), T3~T6各处理均显著高于CK(P< 0.05)。在NaCl+甜菜碱处理下, 与NaCl相比, T1降低了115.23%, T2降低了84.77%, 但T6没有降低, 说明添加甜菜碱能缓解低浓度的盐胁迫伤害, 但对高浓度盐胁迫作用不明显。
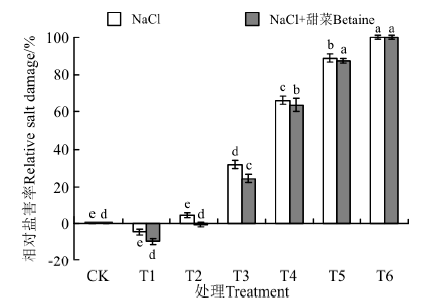 | 图5 不同处理下黑果枸杞种子的相对盐害率Fig.5 Relative salt damage rate of Lycium ruthenium seeds under different treatments |
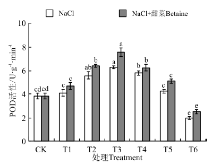
在NaCl处理下(图6), 幼苗中POD含量随浓度的增加而呈现出了先升高后降低的趋势, 在T3时最大, 为6.28 U· g-1, 是CK的165%, T6时最小, 为1.99 U· g-1; 在NaCl+甜菜碱处理下, 幼苗中POD活性均比NaCl处理下的有不同程度地增加, 但也随着处理浓度的增加而逐渐下降; T2~T6各处理均与CK间存在显著性差异(P< 0.05)。其中NaCl+甜菜碱处理下的T3最大, 为7.59 U· g-1, 是NaCl处理下T3的121%; T4~T6, 在NaCl+甜菜碱处理下较NaCl处理, 幼苗中POD活性下降的幅度减小。
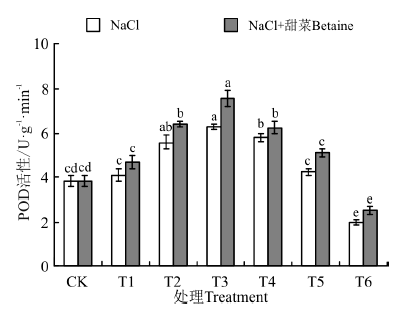 | 图6 不同处理下黑果枸杞幼苗POD的活性Fig.6 POD activity of Lycium ruthenium seedling leaves under different treatments |
在NaCl和NaCl+甜菜碱处理下(图7), 幼苗中SOD活性均随浓度的增加呈现先升高后降低的趋势, 在T3时最大, 分别为9.28、11.59 U· g-1, 分别是CK的292%、364%。在NaCl处理下, T1~T6与CK间均存在显著性差异(P< 0.05), 而在NaCl+甜菜碱处理下, T2~T5与CK间均存在显著性差异; NaCl+甜菜碱处理相比于NaCl处理, 幼苗中SOD含量均出现了不同程度的增加, 且在T4-T6处理间, 随浓度增大而下降, 但下降的幅度较小。
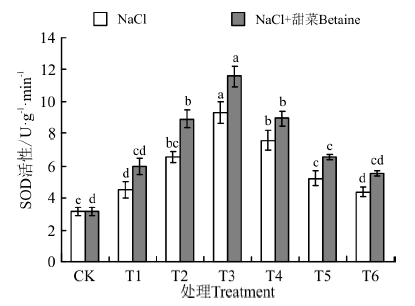 | 图7 不同处理下黑果枸杞幼苗SOD的活性Fig.7 SOD activity of Lycium ruthenium seedling leaves under different treatments |
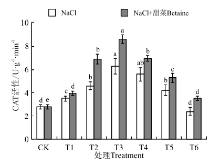
在NaCl和NaCl+甜菜碱处理下(图8), 幼苗中CAT活性均随处理浓度的增加呈现先升高后降低的趋势, 在T3时最大, 分别为6.28、8.59 U· g-1, 分别是CK的224%、307%。在NaCl处理下, T1~T5与CK间均存在显著性差异(P< 0.05), 而在NaCl+甜菜碱处理下, T2~T5与CK间均存在显著性差异; NaCl+甜菜碱相比于NaCl处理, 幼苗中CAT含量均出现了不同程度的增加, 其中最大增加了36.78%; 在T4~T6处理时, CAT活性均随浓度增大而下降, 但NaCl+甜菜碱处理下的下降幅度较小。
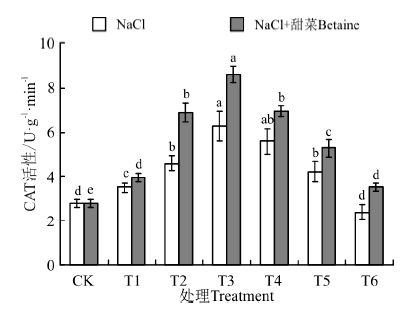 | 图8 不同处理下黑果枸杞幼苗CAT的活性Fig.8 CAT activity of Lycium ruthenium seedling leaves under different treatments |
植物最重要的繁殖器官是种子, 因此植物种子在萌发阶段的耐盐状况可以反映该物种的耐盐能力[14]。通常情况下, 非盐生植物在盐胁迫环境中其生长受到抑制, 而盐生植物生长则需要一定浓度的盐环境才能正常生长, 只有当外界盐浓度超过生长最适盐度时其生长才减缓, 甚至在缺Na+时不能完成其生活史[15]。本研究中, 当NaCl浓度小于25 mmol· L-1时, 对黑果枸杞种子的萌发有一定的促进作用, 但促进作用不明显, 当浓度大于25 mmol· L-1时, 则有抑制作用, 且抑制程度随胁迫浓度的增加而加强, 当浓度达到300 mmol· L-1时, 种子失去萌发能力, 这与其他研究者对黑果枸杞[10, 11]和紫苏(Perilla frutessens)[16]的研究结果一致。在添加外源甜菜碱后, 盐胁迫下种子的发芽率、发芽势、活力指数等萌发指标均有不同程度的增加, 且在随盐浓度增加而下降时, 降低的幅度变小。说明黑果枸杞是盐生植物, 其种子的萌发需要一定的盐分, 适度的盐分可促进种子萌发, 但过高的盐分则会对种子萌发产生抑制作用, 与关于红砂(Reaumuria songarica)[17]的研究结果一致。添加外源甜菜碱可有效的缓解盐胁迫对其种子萌发的伤害作用。
盐胁迫可引起体内产生活性氧簇(ROS), 由于ROS的增加, 细胞体内的抗氧化酶(SOD、POD和CAT)活性都会有所升高以清除产生的ROS[15, 18]。一般来讲, 在盐分胁迫下, 植物体内的(SOD、POD、CAT)等酶活性与植物的抗氧化胁迫能力呈正相关, 能有效地清除活性氧, 抑制膜质过氧化[16]。逆境中保护酶活性增强或维持较高的水平, 才能清除活性氧自由基使之保持较低水平, 防止自由基对生物膜结构和功能的破坏[19]。本研究中, 随着盐胁迫浓度的增加, 抗氧化酶(SOD、POD、CAT)活性均不同程度地表现为“ 先上升, 后下降” 的趋势; 在100 mmol· L-1的范围内, SOD、POD和CAT活性均出现了不同程度的升高, 且均高于CK; 当盐胁迫浓度大于100 mmol· L-1时, 保护酶活性均随浓度的增加而下降; 原因是在低浓度盐胁迫下, 幼苗可通过提高保护酶活性加强耐盐性从而减轻盐对自身的伤害, 但这种能力是很有限的, 当盐含量超出一定程度后, 保护酶的活性就会降低, 这与谭会娟等[20]对红砂耐盐性的研究结果是一致的。在添加外源甜菜碱后, 在各个处理下的SOD、POD和CAT活性均高于NaCl处理下的值, 在盐胁迫浓度大于100 mmol· L-1并不断增大时, 三大保护酶活性均随浓度的增加而下降, 但降低的幅度减小。
甜菜碱作为一种非毒性的渗透调节物质, 主要分布于植物叶绿体和细胞质中逐渐积累以降低渗透势, 同时作为保护物质维持植物正常的生理功能和代谢活动[2], 在盐胁迫环境下, 其还可以不同程度地提高细胞保护酶的活性, 降低活性氧自由基对质膜的伤害和膜脂过氧化作用水平, 维持细胞质膜的稳定性和完整性[21]。相关研究也证明了盐胁迫下施用适宜浓度的甜菜碱可改善黑果枸杞幼苗的耐盐能力, 提高其对盐胁迫逆境的适应性[21]。综上所述, 盐胁迫下添加适宜浓度的外源甜菜碱可有效的缓解盐胁迫对黑果枸杞种子萌发的影响, 提高其萌发率, 萌发速度和出苗整齐度等; 可显著地提高抗氧化酶(SOD、POD、CAT)活性, 增强了清除细胞内氧自由基的能力, 减轻了盐胁迫对其幼苗的伤害, 为黑果枸杞的引种驯化、种质资源的保护以及西北干旱盐碱地的合理利用提供科学数据和理论依据。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|