
{kind=link}
{kind=link}
{kind=link}
放牧和围封对藏北高寒草原紫花针茅群落生物量分配及碳、氮、磷储量的影响
[洪江涛1, 2  , 吴建波
, 吴建波1 , 王小丹1 ]
, 吴建波]
|
|


第一作者:洪江涛(1986-),男,安徽宁国人,在读博士生,主要从事草地生态化学计量方面的研究。 E-mail:hongjiangtao10@163.com
草地围封工程是改善退化草原生态环境和促进牧区经济可持续发展的一项重大举措。以典型高寒草原紫花针茅( Stipa purpurea)群落为研究对象,选取自由放牧、围封4年和围封8年3块样地,比较分析不同草地管理方式下群落生物量分配和碳(C)、氮(N)、磷(P)储量差异。结果表明,自由放牧、围封4年和围封8年的群落地上生物量分别为46.12、146.40和256.44 g·m-2,0-15 cm土层根系生物量分别为274.74、214.87和764.59 g·m-2,15-30 cm土层根系生物量分别为17.80、17.56和31.64 g·m-2;围封显著促进了植被群落的增长( P<0.05),其中对围封4年的优势种紫花针茅和青藏苔草( Carex moorcroftii)总生物量影响最显著( P<0.05)。围封显著提高了植物群落地上部分C、N、P储量,其中围封8年样地营养元素储量最高( P<0.05)。研究结果有助于揭示草地围封工程对藏北高寒草原紫花针茅群落物质分配的影响,为评估该区域退牧还草的生态效应提供了基础数据。
The “grassland enclosure project” is an important measure to improve ecological and environmental quality of degradation grassland and promote economy sustainable development of pastoral area. In the present study, the Stipa purpurea community biomass allocation and carbon(C), nitrogen(N) and phosphorus (P) pools were investigated in three utilization management blocks including free grazing grassland (FG), four years enclosed grassland(F4) and eight years enclosed grassland (F8) on the northern Tibet Plateau alpine. The results showed that aboveground biomass of S. purpurea community in FG, F4 and F8 were 46.12, 146.40 and 256.44 g·m-2, respectively. Root biomasses in layer of 0-15 cm were 274.74, 214.87 and 764.59 g·m-2 and root biomasses in layer of 15-30 cm were 17.80, 17.56 and 31.64 g·m-2 for these three treatments, respectively. These results suggested that enclosure significantly increased ( P<0.05) the community growth. S. purpurea and Carex moorcroftii had the highest biomass in F4 ( P<0.05). Enclosure significantly increased plant C, N and P pools of S. purpurea community and F8 had the highest nutrient pools ( P<0.05) among the three blocks. Our research provide basic data for revealing plant biomass and nutrient allocation response to grassland utilization management and evaluating ecological effects of “grassland enclosure project”.
草地生态系统是我国面积最大的陆地生态系统类型, 总面积达4亿hm2, 约占国土总面积的40%[1]。然而由于长期过度放牧, 草地生态系统结构和功能受到严重的破坏, 对区域生态环境和牧民生产生活均造成巨大的威胁[2]。生物量不仅是反映植物净初级生产力的指标, 而且是评价草地生态系统结构和功能的重要参数之一[3], 它对探讨植物生存策略和生态系统物质循环具有重要的意义[4, 5]。碳(C)、氮(N)和磷(P)是植物必备的大量元素, 是生命体遗传变异和能量代谢等生命过程的重要基础, 近年来也已成为草地生态系统研究的热点及难点[6, 7, 8, 9]。虽然适度放牧可以通过促进植物补偿性生长而提高草地的净初级生产力, 但是草、畜产品的持续输出也会造成矿质元素(C、N和P等)从草地系统中大量流失, 从而导致草地生态系统营养元素循环的失衡[10]。
高寒紫花针茅(Stipa purpurea)草地是西藏自治区主要的草地类型之一, 约占全区草地面积的40%[11]。藏北高寒草地是世界上典型的高寒生态系统, 其优质牧草也为藏北的畜牧业发展提供了良好的条件[12]。据统计, 藏北地区由过度放牧导致草地重度退化面积呈急剧增加趋势, 特别是近20年来, 草地退化面积已达77.1%[13, 14]。为遏制草地退化的趋势, 自治区政府自2004年已开展退牧还草工程[15]。经过若干年的草地围封工程的实施, 群落的恢复演替状况如何?优势物种和伴生种生长状况如何?植物营养元素特征发生了怎样的变化?这些问题还缺乏基础数据去评估。本研究以高寒草原紫花针茅群落为研究对象, 采集自由放牧和不同围封年限样地的植物样品, 分析放牧和围封样地间植物群落生物量和C、N、P生态化学计量学特征及其储量的差异性, 阐明不同草地管理方式对高寒紫花针茅群落植物生长和营养元素状况的影响, 以期为进一步有效地实施退牧还草工程提供理论和实践依据。
研究地点位于西藏自治区那曲地区安多县。该区域属高原亚寒带半湿润季风气候区, 空气稀薄, 昼夜温差大, 无绝对无霜期, 多风雪天气。年日照时数为2 847 h, 年降水量约为435 mm。年平均气温-2.8 ℃, 7月份平均气温为7.8 ℃, 1月份平均气温为-14.6 ℃, 年大风日数在200 d以上。
样品采集于那曲地区安多县措玛乡(2012年8月)。选择围封年限分别为4年(2008年围封)和8年(2004年围封)的紫花针茅群落以及1块自由放牧的紫花针茅群落样地为试验地点。3块试验场地形和土壤条件一致, 并且均位于地势平坦的高原面上, 相距1 km之内, 每块样地面积约为7 000 m2。建群种均为紫花针茅和青藏苔草(Carex moorcroftii), 主要伴生种有矮火绒草(Leontopodium nanum)、冰川棘豆(Oxytropis glacialis)、二裂委陵菜(Potentilla bifurca)等。
在3块试验场地随机设置一个10 m× 10 m的大样方, 在样方内随机设置3个1 m× 1 m的小样方, 取植物群落样品。群落植物样品采集包括群落地上生物量和根系生物量。齐地刈割群落地上部分并装入编号信封内。然后用铁锹挖出根系0-15和15-30 cm土柱(含根), 装入布袋带回实验室后用0.178 mm筛子冲洗和分离, 取得根系部分生物量。根据根的韧性和颜色去除死根[16]。然后在每个大试验场内用样线法随机采集10株优势物种紫花针茅和青藏苔草以及主要伴生种矮火绒草和冰川棘豆。采集方法为土柱法, 即用铁锹铲将包含目标物种长宽深分别为15 cm× 15 cm× 30 cm的土柱取出, 剔除根系表面上的泥土, 保证每株植物的完整性。所有物种样品清洗干净后晒干带回实验室, 装进纸信封晾干。
植物样品(包括群落和物种)105 ℃下杀青15 min后, 在烘箱内(65 ℃)烘干至恒重, 测定其生物量(精确到0.01 g)。然后群落植物样品分植物器官装袋, 研磨过直径0.25 mm筛子, 待测理化性质。植物群落C含量采用重铬酸钾容量法测定, 全N测定采用凯氏定氮法, 全P测定采用钼锑抗比色法[17]。
群落植被养分储量由单位面积内群落生物量和营养元素含量相乘估算。用单因素方差分析(ANOVA)检验比较自由放牧、围封4年和围封8年间紫花针茅植物群落和物种生物量分配和营养元素特征的差异性。用Duncan多重比较法进行显著性检验(LSD)。数据处理均在SPSS 16.0中进行, 图形绘制由Sigmaplot 11.0绘制。
自由放牧、围封4年和围封8年的群落地上生物量分别为46.12、146.40和256.44 g· m-2。与自由放牧相比, 围封显著提高了紫花针茅群落地上生物量(P< 0.05), 并随着围封年限的增长, 对生物量促进作用更明显。围封8年地上生物量显著高于围封4年和自由放牧样地(P< 0.05)(图1)。
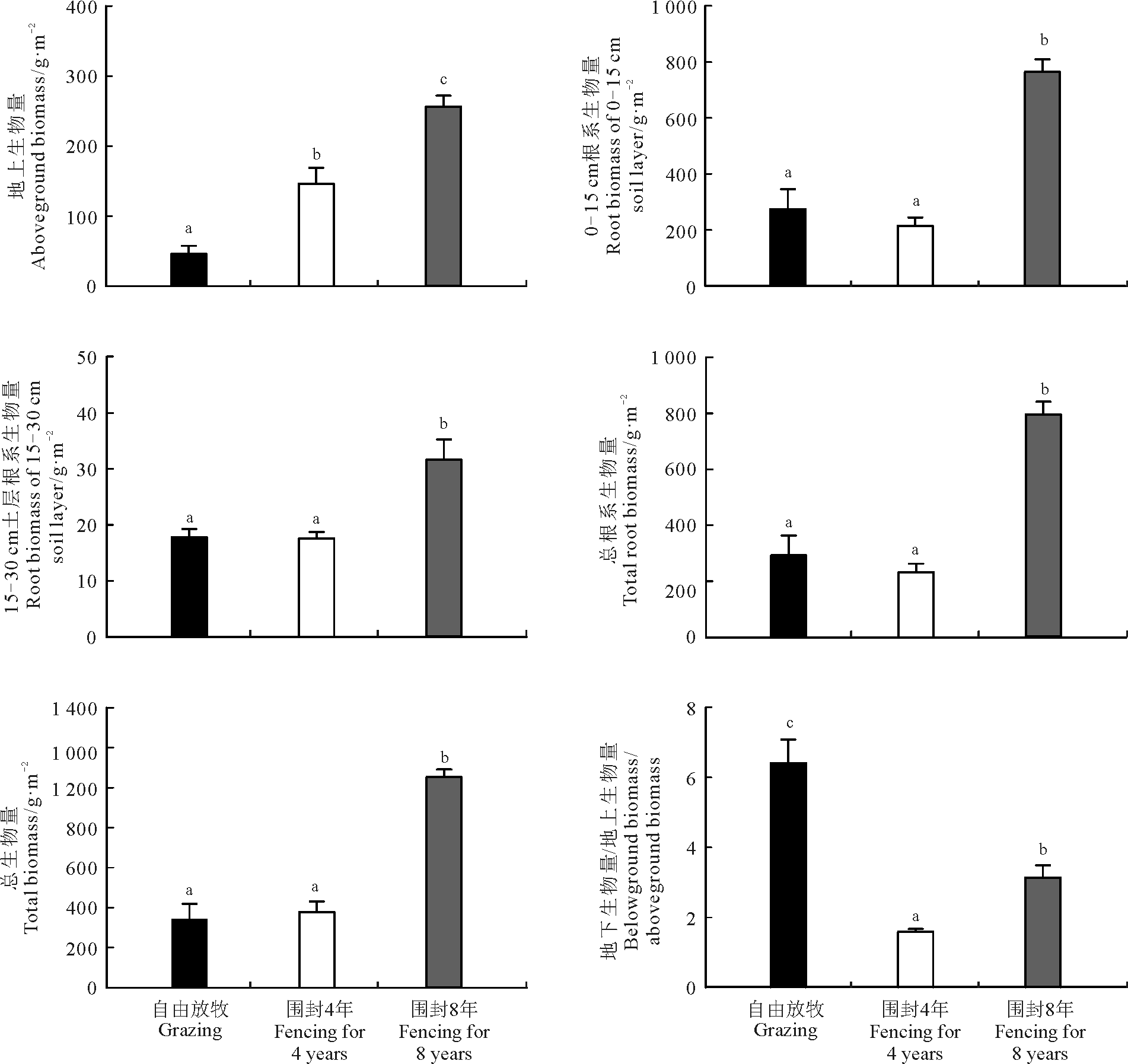 | 图1 自由放牧和不同围封年限高寒紫花针茅群落生物量分配注:图中不同小写字母表示不同样地间紫花针茅群落各组分生物量分配显著差异(P< 0.05)。图2同。Fig.1 Biomass allocation of Stipa purpurea community in grazed and fenced grasslandsNote: Different lower case letters represent significant difference in biomass allocation of Stipa purpurea community among different grasslands at 0.05 level. The same in Fig.2. |
围封4年与自由放牧样地的0-15和15-30 cm根系生物量以及根系总生物量均无显著差异(P> 0.05)(图1)。围封8年样地则具有最高的根系生物量(P< 0.05)。与自由放牧样地相比, 围封4年总生物量虽然呈轻微增加趋势, 但是并未达到显著水平(P> 0.05), 而围封8年在不同利用方式中具有最高的总生物量(P< 0.05)(图1)。
自由放牧、围封4年和围封8年地下生物量和地上生物量的比值 (Mb/Ma) 分别为6.41、1.59和3.14。围封显著降低了紫花针茅群落Mb/Ma值, 且围封4年样地具有最低的Mb/Ma值(P< 0.05)(图1)。
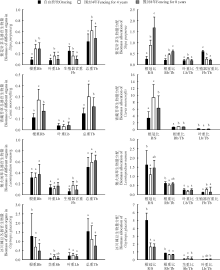
从单个物种来看, 围封显著提高了紫花针茅的根重, 但是围封4年的叶重和总重值最高(P< 0.05)(图2)。随着围封年限的增长, 根冠比(R/S)值和根重比(Rb/Tb)逐渐增大, 生殖器官重比(Fb/Tb)逐渐减小(P< 0.05)。其中在围封4年样地紫花针茅分配给植物叶片的生物量比例最高(P< 0.05)(图2)。
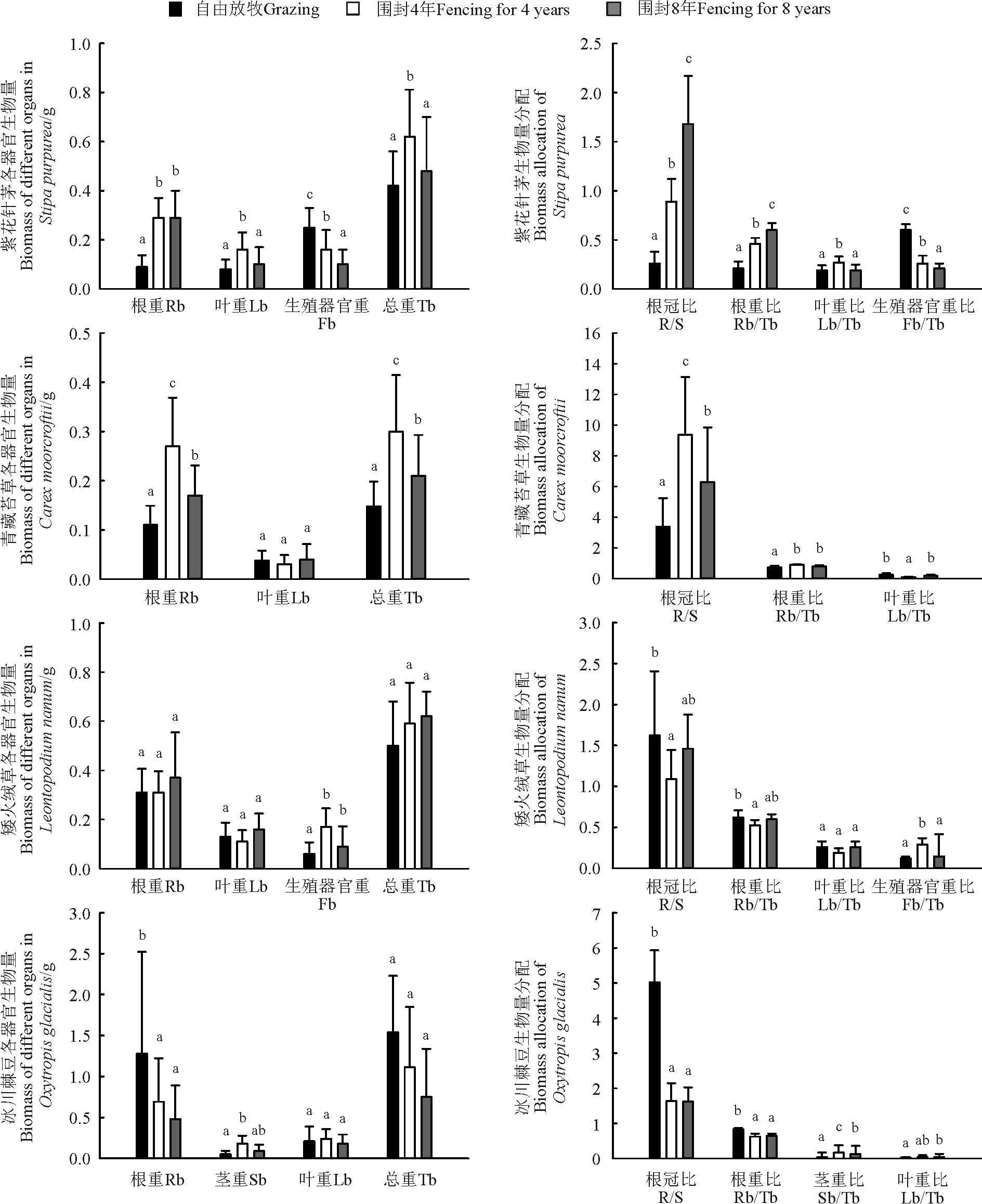 | 图2 自由放牧和不同围封年限样地主要物种生物量分配注:Rb-根重, Sb-茎重, Lb-叶重, Fb-生殖器官重, Tb-总重, R/S-根冠比, R/T-根重比, L/T-叶重比, S/T-茎重比, F/T-生殖器官重比。Fig.2 Biomass allocation of main species in grazed and fenced grasslandsNote: Rb-Root biomass, Sb-Stem biomass, Lb-Leaf biomass, Fb-Flower biomass, Tb-Total biomass, R/S-Root biomass/shoot biomass, Rb/Tb-Root biomass/total biomass, Lb/Tb-Leaf biomass/total biomass, Sb/Tb-Stem biomass/total biomass, Fb/Tb-Flower biomass/ total biomass. |
围封显著提高了青藏苔草的根重(P< 0.05), 但是叶重变化不显著(P> 0.05), 围封4年青藏苔草总重最高(P< 0.05)(图2)。围封4年具有最高的R/S(P< 0.05), 自由放牧的Rb/Tb值最低(P< 0.05)(图2)。
围封对矮火绒草根重、叶重和总重影响不显著(P> 0.05), 但围封提高了其生殖器官的生物量, 围封4年的生殖器官重值最高(P< 0.05)(图2)。围封4年样地的R/S和Rb/Tb值最低(P< 0.05), 而Fb/Tb值最高(P< 0.05)(图2)。
围封显著降低了冰川棘豆的根重(P< 0.05), 但是对叶重和总重影响不显著(P> 0.05)(图2)。在3块样地中, 自由放牧具有最高的R/S和Rb/Tb值(P< 0.05)(图2)。
自由放牧、围封4年和围封8年植被地上部分的C含量和P含量均没有明显差异(P> 0.05), 而自由放牧群落地上部分N含量(14.54 mg· g-1)显著高于围封4年(12.37 mg· g-1)和围封8年地上部分N含量(11.96 mg· g-1)(P< 0.05)(表1)。
| 表1 自由放牧和不同围封年限紫花针茅群落各部分C:N :P生态化学计量学特征 Table 1 C:N :P stoichiometry of Stipa purpurea community in grazed and fenced grasslands |
自由放牧、围封4年和围封8年0-15 cm土层根系的C含量和N含量均没有明显差异(P> 0.05)。自由放牧群落0-15 cm土层根系P含量(0.24 mg· g-1)显著小于围封4年(0.51 mg· g-1)和围封8年样地(0.42 mg· g-1)(P< 0.05)的(表1)。
自由放牧、围封4年和围封8年15-30 cm土层根系C含量没有明显差异(P> 0.05), 围封4年15-30 cm土层根系N(7.11 mg· g-1)含量显著小于自由放牧(12.71 mg· g-1)和围封8年样地(13.16mg· g-1)(P< 0.05)。自由放牧样地15-30cm土层根系P含量显著高于围封4年(P< 0.05), 而围封8年15-30 cm土层根系P含量与自由放牧和围封4年样地的没有显著差异(P> 0.05)(表1)。自由放牧样地地上生物量的N:P最高(19.54), 围封4年样地N:P最低(14.18)(P< 0.05)。且自由放牧样地0-15 cm土层根系生物量具有最高的N:P值(53.27)(P< 0.05)(表1)。
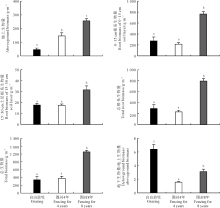
自由放牧、围封4年和围封8年的群落地上部分C储量分别为20.03、65.66和111.44 g· m-2, N储量分别为0.57、1.84和3.75 g· m-2, P储量分别为0.03、0.13和0.19 g· m-2。围封显著提高了植物群落地上部分C、N和P储量(P< 0.05), 且随着围封年限的增长, 营养元素储量越高(图3)。自由放牧和围封4年样地0-15 cm土层根系植物C、N和P储量没有显著差异(P> 0.05), 而围封8年0-15 cm土层根系生物量C、N和P储量最高(P< 0.05)。这3块样地0-15 cm土层根系生物量P储量没有显著差异(P> 0.05)。围封8年的15-30 cm土层根系具有相对较高的C和N储量(P< 0.05)。而自由放牧和围封4年样地15-30 cm土层根系生物量的C、N和P储量则没有显著差异(P> 0.05)。
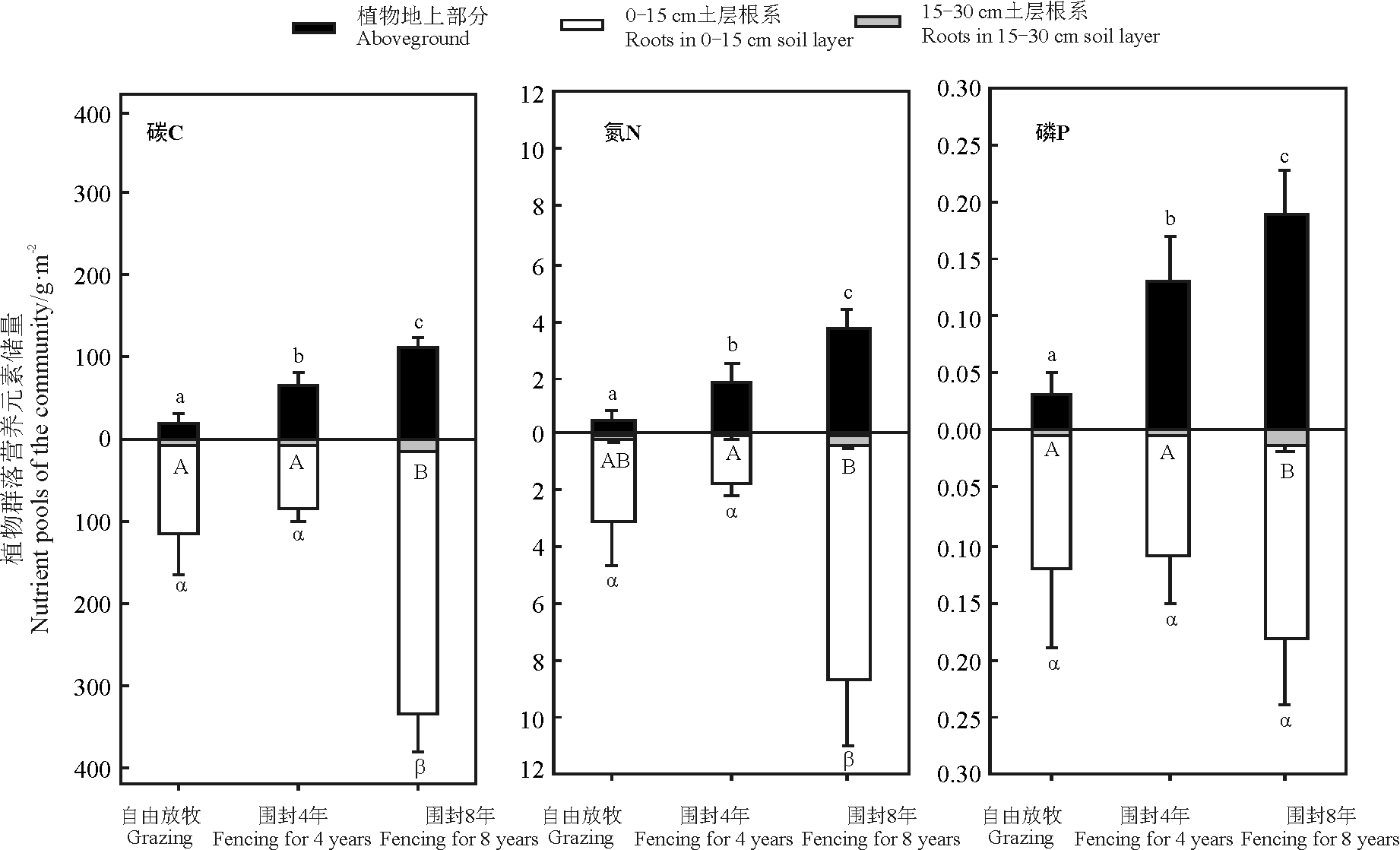 | 图3 自由放牧和不同围封年限高寒紫花针茅群落植物C、N和P储量注:不同字母表示自由放牧和不同围封年限3个样地群落不同组分的C、N和P储量差异性(P< 0.05)。Fig.3 Carbon, nitrogen and phosphorus pools of Stipa purpurea community in grazed and fenced grasslandsNote: Different letter represented significant difference in carbon, nitrogen and phosphorus pools at 0.05 level in grazed and fenced grasslands. |
放牧和围封作为不同的草地管理方式, 通过动物的采食、践踏和排泄物归还等对草原生态系统造成不同程度的影响, 主要表现在草原植物生产力、种子萌发、物质分配、物种多样性和群落结构等方面[15, 18, 19]。自由放牧样地由于牦牛(Bos mutus)和藏绵羊(Ovis aries)长期对地上部的啃食作用, 使得其地上生物量比围封草地低, 根冠比最高, 而退化草地(自由放牧)围封后, 原来由于遭受抑制生长的植物得以休养生息, 促进了其幼苗的萌发、分蘖数的增加, 最终提高了草地的生产力[20]。围封对紫花针茅群落总生物量有显著的恢复作用, 并随着围封年限的增长, 恢复程度越好。但是我们注意到, 围封4年与自由放牧样地根系生物量没有显著差别, 而地上生物量呈显著增加趋势。这说明短期围封后(4年), 动物采食压力得以解除, 群落优先分配生物量在地上部分, 植物地上生物量快速增加, 造成其根冠比最低。
通过对比分析优势种紫花针茅和青藏苔草在3块样地的生物量及其分配特征, 发现围封4年已经使得这两个物种根重和总重达到最大值, 说明围封4年已经使得优势物种的生物量得到明显的恢复, 甚至高于围封8年样地。可能原因是围封年限过长(8年)导致植物密度增大, 限制了牧草在放牧条件下的超补偿生长机制作用[21, 22], 降低植物生产的周转率[23], 从而影响到物种对资源的利用效率[24]。由此可见, 就高寒紫花针茅群落恢复时间尺度而言, 围封4年已经显著提高了优质牧草紫花针茅和青藏苔草的产草量, 这对指导草地恢复有非常重要的意义。但是由于缺乏对植物物种C、N和P特征的分析, 目前还无法建立生物量分配与物种养分功能性状的相关关系, 这方面内容还有待于今后进一步研究。
C、N和P不仅是植物体内的基本营养元素, 而且是参与植物-土壤生物地球化学循环的重要物质。草地管理方式对植物C含量的影响不显著, 主要是由于植物体内的C一般不会直接参与植物的生产, 其在植物体内起着骨架的作用, 具有很小的变异性。此外, 家畜的不断采食, 可能会促进营养物质向地上幼嫩器官重新分配, 从而使得自由放牧草地植物地上活体具有较高的N含量。这与割草促进了内蒙典型温带草原植物N素增加的研究[25]结果相类似。比较自由放牧、围封4年和围封8年植物的N:P数值, 可以反映3块样地营养元素的相对受限状况[26, 27]。例如:自由放牧与围封样地相比, 其地上部和0-15 cm土层根系植物组织的N:P较高, 可以反映出自由放牧样地相对受P素限制; 而围封4年N:P值相对较低, 表明其相对受N素限制, 主要原因是围封4年正处于恢复演替的过程, 地上部分生物量急剧积累, 需要有大量的养分需求(特别是N素)。
对紫花针茅群落地上和根系部分C、N和P储量的分析发现, 随着围封年限的增长, 群落地上部分的C、N和P储量均逐步增加。说明围封后, 解除了牲畜对牧草的采食压力, 植物地上部分生物量得以迅速的恢复, C、N和P营养元素库也得到显著增加。但是围封4年样地和自由放牧样地0-15和15-30 cm土层根系生物量以及C、N和P储量没有差异性, 说明高寒草原在恢复初期对群落根系营养元素库促进作用不明显, 而是投入更多的生物量和营养物质到地上部分促进群落光合作用。草地围封8年后, 其地上、根系和总营养元素恢复状态是3个样地中最佳的。由于放牧和围封样地植物C、N和P营养元素含量差别较生物量差异小, 因此, 植物群落C、N和P储量差异主要受3个样地生物量的影响, 与董晓玉等[10]在黄土高原地区草地围封试验的结果较为一致。本研究中, 自由放牧、围封4年和围封8年地上部分总C储量分别为20.03、65.66和111.44 g· m-2, 自由放牧样地群落地上部分C储量比Fan等[28]研究报道的高寒草原的地上C储量(31 g· m-2)结果低, 而围封样地C储量明显高于其研究结果。本研究中自由放牧、围封4年和围封8年根系部分总C储量分别为121.96、90.72和347.73 g· m-2, 明显低于Fan等[28]研究报道的根系总C储量的值(1 226 g· m-2)。Lu等[29]研究报道的典型紫花针茅草原地上部分N储量为2.60 g· m-2, 介于本研究围封4年和围封8年样地之间。与前人研究结果存在出入, 主要是由于不同研究地点群落生物量的差异造成的, 而植物营养元素含量差异不明显。
在对藏北高寒紫花针茅群落放牧和围封样地的研究发现, 与自由放牧样地相比, 围封显著促进了群落地上部分生物量的增加, 并且随着围封年限的增长生物量累积的更为明显。但是围封4年并没有促进根系生物量显著提高。主要优势物种紫花针茅和青藏苔草在围封4年样地生物量最高。自由放牧样地植物N:P最高, 而在围封4年样地植物N:P相对较低, 表明自由放牧样地相对缺乏P素, 而围封4年样地缺乏N素。围封显著提高了植物群落的C、N和P储量。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|