

{kind=link}
{kind=link}
甘南夏河高寒草甸高原鼢鼠和高原鼠兔秋季栖息地特征
[许国成 ]
]
]
|
|


2010年至2014年连续5年对甘南夏河高寒草甸高原鼢鼠( Eospalax bailyei)和高原鼠兔( Ochotona curzoniae)的分布与其地形、植被因子等进行了调查,分析两小型哺乳动物秋季越冬前的栖息地利用特征。结果表明,其秋季栖息地利用与地形和植被特征显著相关。高原鼠兔易选择坡地、半阳坡和坡中位的地形,而高原鼢鼠易选择平地、阳坡和坡底位的地形;植被盖度是影响高原鼠兔分布的重要因子。地上生物量非中等的平地是两小哺乳动物共存的主要分布地,两小型哺乳动物的种群动态与植被盖度和生物量均显著相关( P<0.05);鹅绒委陵菜( Potentilla anserina)、花苜蓿( Melissilus ruthenicus)、柴胡( Radix bupleuri)和美丽风毛菊( Saussurea pulchra)4种植物多度与两小型哺乳动物的分布显著相关。
A 5-year research was conducted on the features of vegetation and topographic factors of habitats of two dominate small mammal species Eospalax bailyei and Ochotona curzoniae in alpine meadow of Xiahe, Gannan. Significant correlations were found between habitat utilization of these two species and vegetation, as well as geographic factors. Plateau pikas preferred sunny middle slope, while plateau zokors preferred flat and sunny lower slope. The coverage was an important factor predicting the presence of plateau pika. Areas with high or low biomass in flat area were more likely to be occupied by both species. Their coexistence last about 3 years. The abundance of four plant species Potentilla anserine, Melissilus ruthenicus, Radix bupleuri and Saussurea pulchra significantly affected habitat selection by both species.
动物与其生存环境之间的关系一直是生态学研究的经典内容, 两者在相互作用下发挥着重要的生态和进化功能[1]。小型哺乳动物是草地生态系统中固有的成员, 肩负有能量流动、物质循环等作用, 一度作为高原生态系统的指示种[2, 3], 其数量大小和生态功能对生态系统的平衡和稳定具有重要意义[4, 5, 6]。
高原鼢鼠(Eospalax bailyei)和高原鼠兔(Ochotona curzoniae)是青藏高原高寒地区两类主要的优势小哺乳动物[6], 他们分布广泛, 局部地区密度高、数量大, 已成为草原退化的重要因素之一[7], 对其有效防治一直是研究的重要内容[8]。哺乳类(包括啮齿类)等有害生物的防控因化学防治等产生环境污染、抗性种群等问题后[9], 人们一直在探索其他有效的控制和管理方式。生态防控概念的提出引发了人们诸多的思考与实践[10]。基于自然制约力控制草原鼠类数量和分布的生态调控法, 是当今草原鼠害防治发展的主流方向, 也是鼠害防控研究的热点领域[11, 12, 13]。草地生态系统中小型哺乳类和所处环境之间的关系认识, 是寻求生态防控的关键途径之一, 各学者已从不同的方面进行了诸多的研究[14, 15, 16, 17, 18, 19, 20, 21]。从植被特征入手, 分析其与栖息环境的关系, 对于草地生态系统的管理具有重要的参考意义, 可为小哺乳类的生态防控和草地恢复重建提供依据[22, 23]。
夏河县是甘肃省主要的鼠害测报点之一, 也是甘肃省主要的草原鼠害发生区域[24]。高原鼢鼠和高原鼠兔是该县主要的危害小哺乳动物。而秋季是该两类小哺乳动物越冬的关键时期, 影响其存活和来年的繁殖等。秋季也是一年当中第2个主要危害时期, 该时期是有效防控的关键时机, 探讨其与栖息环境的关系至关重要。本研究基于连续5年的监测数据, 分析高原鼠兔、高原鼢鼠及其两哺乳类同域共存下其分布格局与地形因子、植被特征等的关系, 并探讨两类物种共存和单个存在下草地植被特征等, 旨在为草地生态系统中动植物相互关系研究、小哺乳类动物资源的管理及促进草地恢复与重建等提供理论依据。
夏河县位于101° 54'-103° 25' E、34° 32'-35° 34' N。地处青藏高原东北部边缘, 地形为山原地貌, 地势高、地形复杂。气候属寒冷湿润类型, 高原大陆性气候特点比较明显。土壤包括高山草甸土、亚高山草甸土、亚高山草原土、草甸土和亚高山草原土, 另有暗棕壤、沼泽土和栗钙土镶嵌分布。海拔为2 900~4 600 m, 年平均气温2.6 ℃, 年均降水量516 mm, 年均无霜期56 d, 年日照时间为2 296 h。境内草地总面积为753.87万亩(约合52.258万hm2), 牧草种类72科290属628种, 可食牧草574种, 占牧草种类的91.4%, 其中优良牧草43种。优势植物主要有披碱草(Elymus dahuricus)、早熟禾(Poa annua)、矮嵩草(Kobresia humilis)、苔草(Carex tristachya)、艹洽草(Koeleria glauca)等, 是甘南州重要的牧业基地之一。
选择夏河鼠害的主要分布区域, 排除旅游等人类干扰的影响, 选择放牧和其他情况较一致的区域, 通过定点选择30个样区, 样区面积1~2 hm2, 于2010-2014年的每年9月下旬, 样区内随机选取3个面积为20 m× 50 m的样地, 采用堵洞盗洞法测定试验样地内高原鼠兔有效洞口数, 每天09:00-10:00 测定一次, 连续测定3 d。同时用样方捕尽法, 连捕3 d统计高原鼢鼠的数量。在2010年秋季记录每个样地的高原鼢鼠和高原鼠兔分布情况, 此后尽量在同一样区做调查, 来分析两种动物共存时间。若同一样区高原鼢鼠和高原鼠兔迁入情况等较大, 就选择临近样区做调查, 据此判断两物种的密度大小和共存时间。样方捕尽法调查高原鼢鼠的种群密度时, 采用不同的样区数据进行, 各样区之间相距较远, 以保证种群数量的客观性。
根据该地区的地形地貌, 将调查样地坡位分成3个等级, 平地、坡地和沟谷。把样方所处地的坡位分为4 级, 即坡上位(山岗和坡上部)、坡中位(山腰和坡中部)和坡下位(坡下部)和坡底位(谷底)。将样地坡向划分为阴坡、半阴坡、半阳坡和阳坡4 种坡向, 即阴坡:0~45° 和315° ~ 360° ; 半阴坡:45° ~90° 和 270° ~315° ; 半阳坡:90° ~135° 和225° ~270° ; 阳坡:135° ~225° 。
同时每样地随机取1 m× 1 m的3个样方, 测定植被因子, 即多度(每个样方的前5个优势种)、盖度、地上生物量(干重)3项指标。多度采用Braun-Blanquet多度等级, 多度和盖度采用目测法估计, 每样方3次, 植物群落地上部分生物量的测定采用分种齐地面刈割, 风干后称重。
采用SPSS 19.0软件对所测数据统计及多元回归分析, 用平均值和标准误表示测定结果, 分别对样方间植被盖度、生物量和种群密度等进行单因素方差分析, 并用Duncan法对各测定数据进行多重比较; 采用Excel 2003制图。
根据调查的结果, 本研究发现啮齿类的分布格局受地形因子的影响较大, 但地形因子对啮齿类分布的影响不同, 地形因子影响下的环境条件是啮齿类分布的主要因素。在高原鼢鼠80个样方中, 坡地有17个, 平地63个(表1)。坡地中4个为半阴坡, 其余为阳坡和半阳坡, 坡位除2个在坡中位外, 其余均在坡底部; 高原鼠兔30个样方中, 17个为坡地, 10个沟谷, 3个平地, 坡向间差别不大。坡位显示, 12个为坡中位, 5个坡下位; 而在同域共存的分布区35个样方中, 平地有29个, 坡地6个, 但全选择半阳坡生境, 坡底位和坡中位各有3个。夏河高寒草甸啮齿类冬季的分布显示出一定的地形偏好性和规律。
| 表1 啮齿类分布区地形因子指标频数个样方 Table 1 Frequency distribution of topographic factors with different rodent speciesplot |
研究发现, 啮齿类分布格局与植被特征有极大关系。一是植被多度与啮齿类的分布明显有关。除了当地优良牧草披碱草、早熟禾等外, 与高原鼢鼠相伴而生的3个优势植物是鹅绒委陵菜(Potentilla anserina)(占调查样方的43.8%)、花苜蓿(Melissilus ruthenicus)(占调查样方的50.0%)、柴胡(Radix bupleuri)(占调查样方的37.5%); 与高原鼠兔相伴而生的优势植物是鹅绒委陵菜(占调查样方的93.3%), 美丽风毛菊(Saussurea pulchra)仅占调查样方的6.7%; 而在高原鼢鼠和高原鼠兔同域共存的分布地, 鹅绒委陵菜(占调查样方的93.0%)、翻白委陵菜(P. discolor)(占调查样方的7.0%)和美丽风毛菊(占调查样方的7.0%)是优势植物, 这些优势植物的多度均在较多级及25%以上。二是植被盖度与啮齿类的分布格局关系较大, 盖度较高的样地, 啮齿动物种群数量小, 相反, 盖度较低的样地, 啮齿类种群数量大。三是啮齿类的分布和地上生物量也有一定的关系(表2)。地上生物量与啮齿类种群密度的关系也出现了明显的规律, 地上生物量偏高的样地, 两类啮齿类的密度均较高。而地上生物量非中等的样地是两种啮齿类共存的主要分布地, 在生物量偏低和偏高的样地, 两种啮齿类共存较多。
| 表2 小哺乳类不同密度区植被两项指标频数分布只 Table 2 Frequency distribution of vegetation index with different small mammals densitieshead |
高原鼢鼠栖息地之间的植被盖度除2010年, 2011年和2012年间差异不显著外, 均和2013年, 2014年间达到显著水平(P< 0.05)。但地上生物量在各年份间差异不显著(图1); 高原鼠兔各栖息地之间, 植被盖度和地上生物量在年份间的差异均不显著(图2)。为确定植被盖度和地上生物量对两啮齿类种群动态的综合影响, 本研究又进行了多元回归分析。研究发现, 高原鼢鼠的种群密度(Y)和地上生物量(X1)与植被盖度(X2)之间显著相关, 其回归方程为:Y=190.7-0.086X1-0.379X2(R2=0.996, P< 0.01)(图1); 高原鼠兔也呈显著相关关系, 其回归方程为:Y=560.6-4.026X1-0.533X2, (R2=0.898, P< 0.01)(图2)。
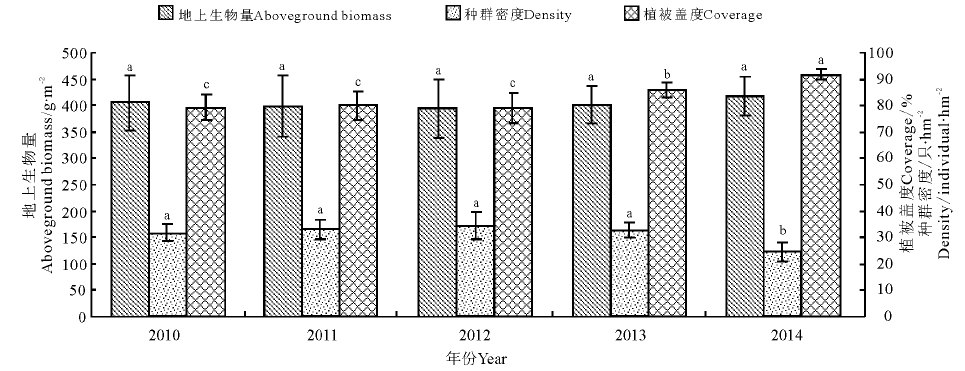 | 图1 高原鼢鼠种群密度和植被盖度及生物量的关系注:不同小写字母表示同一指标不同年份间差异显著(P< 0.05)。下同。Fig.1 Relationships between coverage, aboveground biomass and population dynamics of plateau zokorNote: Different lower case letters for the same parameter show significant difference among different years at 0.05 level. The same below. |
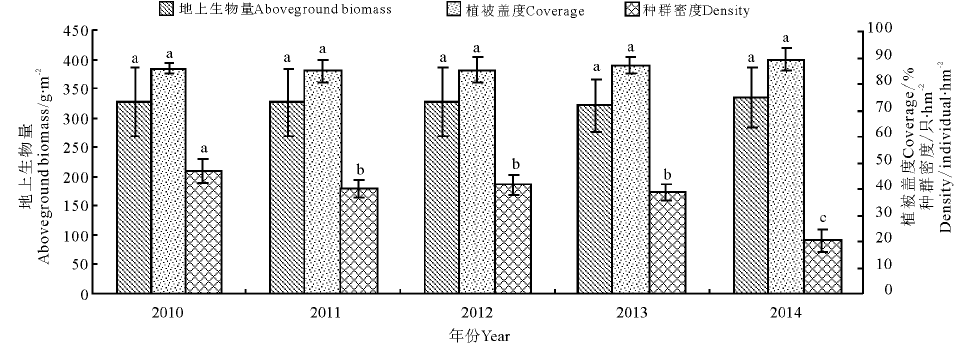 | 图2 高原鼠兔种群密度和植被盖度及生物量的关系Fig.2 Relationships between plant coverage, aboveground biomass and population dynamics of plateau pika |
高原鼢鼠种群密度2014年是5年中最低的年份, 和其余年份之间差异显著(P< 0.05)(图1); 高原鼠兔种群密度2010年显著高于其他年份, 2014年显著低于其他年份, 2011、2012、2013年年间差异不显著(图2)。在研究每年选定的30个样区中, 检测了小哺乳类分布和共存的情况, 分析可能的相互作用及其分布格局。研究发现, 高原鼢鼠种群动态较为稳定, 在5年的观测中发现了6个样点, 高原鼠兔有5个样方, 而混合存在的样方中只有两个样方有5年的共存时间, 大部分(8个样方)只有3年的共存时间, 占到了调查样方的2/3。两年共存的有两个, 未发现4年共存的样方。
动物对环境的利用是其对各因子的综合适应, 是生存对策的一个权衡[25]。本研究发现, 地形因子对两类小型哺乳动物的影响不同。对于地下生活的高原鼢鼠而言, 更易选择平地、阳坡和坡底位的地形, 推测地形因子下的土壤水分、温度是影响其分布的主要因素, 地形与大气温湿度等交互作用, 影响了土壤的水分、温湿度。而土壤的水分和温度等影响高原鼢鼠的具体活动规律, 地下水分对挖掘活动影响很大, 水分缺少等使土壤的硬度等发生变化, 导致动物挖掘付出极大的能量代价[26]。而温度与其能量的分配等密切相关, 过低的温度会限制其正常的生命活动。对于地上生活的高原鼠兔, 其更易选择在坡地、半阳坡和坡中位的地形, 高原鼠兔地面上的生活区别于高原鼢鼠, 推测与地面活动的风险大小等有关, 地面生活的主动性, 捕食风险是主要的因素[27]。卫万荣等[28]研究了高原鼠兔的洞穴结构, 发现高原鼠兔的洞系建筑特点除了常规意义上的栖居和避险等基本功能外, 还兼顾了洞内温度、对流和抵御寒风的利弊权衡。坡中位的半阳坡可以显著提高土壤温度。这也是地形因子对高原鼠兔影响的综合反映。这些适应特征都说明在能量和生存风险回避等方面, 小型哺乳类动物均有一定的权衡, 是对当地环境的一种适应。
本研究中, 高原鼢鼠的密度调查采用了样方捕尽法, 此法虽较费力, 但能详细地了解其种群的绝对密度, 相较以往的土丘系数法更精确。但该方法也有一定的缺陷, 会干扰其真实的种群密度及其动态。在密度调查完后, 样地的选择上, 尽量加大了相邻样地之间的距离, 以减少鼠个体数量的减小给其种群密度等带来的影响。高原鼢鼠样方捕尽后, 选择同样样地也有可能造成植物群落的差异, 草地的异质性会影响植被特征和高原鼢鼠种群密度的关系。因此, 在样地的选择上通过距离避免了可能的误差。在监测实践中, 细致的数据会加大监测工作的任务, 本研究中对于地下生活的高原鼢鼠, 地下生物量的多少也会极大影响其种群密度。高原鼢鼠地下生活特点为研究带来了不便, 其种群密度的调查一度是个难点, 而影响种群密度的因素等也会因调查方法而产生变化, 后续有望通过进一步优化其调查方法。
植被条件是制约啮齿类种群数量消长和分布的重要因素之一[29]。本研究发现, 植被特征对两类小型哺乳类动物分布均有影响, 但影响不同, 各自有主导因子。对于地面上的高原鼠兔, 植被盖度显著影响其分布, 分析原因可能是盖度的增大一是不利于活动、行走及其逃避等; 二是盖度的下降, 形成了开阔的生境, 有利于躲避敌害[30]。但对于地下生活的高原鼢鼠来说, 地上的盖度和生物量多为植被的综合指标, 显著影响其分布, 相反, 地面盖度下降后, 有利于肥大根系等高原鼢鼠喜食植物的入侵[30], 为其提供了食物资源。张军等[31]发现植被盖度和高度与高原鼢鼠种群密度表现为极显著线性相关, 盖度和高度反映了植被的基本状况, 二者的下降为高原鼢鼠喜食植物的入侵提供了条件, 也为高原鼢鼠的栖息地利用提供了机会。植被的地下生物量是影响高原鼢鼠的主要因素[23], 因在监测实践中工作的不便, 没有监测地下生物量的情况, 后续的研究有必要对地下生物量进行监测。
地上地下生物的作用过程及其格局是群落演替的研究热点[32], 高原鼠兔和高原鼢鼠地上地下的影响机理至今认识不足。张卫国和江小蕾[33]研究了高原鼢鼠和高原鼠兔的共存格局, 发现高原鼢鼠的种群密度随高原鼠兔种群密度的增加而下降; 两种鼠在同一生境下的共栖可以认为是高原鼠兔对高原鼢鼠栖息地选择性迁入的结果; 高原鼠兔与高原鼢鼠的共栖只是一个种间竞争过程的表象, 大约持续3年时间。本研究发现, 两小型哺乳类动物的共存时间多在3年左右, 鲜见4年以上的共存时间。后续的种间关系研究需要更多的关注, 进一步利用其它学科理论和技术分析两者之间的关系, 为草地生态系统食物网、生态链等研究提供依据。
甘南夏河高寒草甸两小型哺乳动物的栖息地利用与地形和植被特征显著相关, 但地形因子对其的影响不同。高原鼠兔更易选择在坡地、半阳坡和坡中位的地形。高原鼢鼠易选择平地、阳坡和坡底位的地形; 植被盖度是影响高原鼠兔分布的重要因子, 但对地下生活的高原鼢鼠影响不显著。地上生物量非中等的样地是两小型哺乳类动物共存的主要分布地, 其种群密度与植被盖度和生物量均显著相关; 鹅绒委陵菜、花苜蓿、柴胡和美丽风毛菊4种植物多度与两小型哺乳类动物的分布显著相关。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|