
{kind=link}
{kind=link}
{kind=link}
{kind=link}
长江源区高寒草地土壤有机质和氮磷含量的分布特征
[高洋1, 2  , 王根绪
, 王根绪1 , 高永恒1 ]
, 王根绪, 高永恒|
|


第一作者:高洋(1984-),女,黑龙江大庆人,博士, 主要从事冻土生态方面的研究。E-mail:35222461@qq.com
土壤养分是高寒草地生态系统维系健康的基础。本研究分析了长江源区沼泽化草甸、典型草甸、草原化草甸和沙生草地的土壤养分特征,以期为高寒草地合理管理提供理论依据。研究表明,不同类型高寒草地的土壤有机质和全氮含量存在明显的差异,表现为沼泽化草甸>典型草甸>草原化草甸>沙生草地,土壤有机质和全氮含量在各个草地类型内均随着土层深度增加而显著降低( P<0.05);土壤速效氮含量为草原化草甸>典型草甸>沼泽化草甸>沙生草地,草原化草甸含量高达65.5 mg·kg-1,而沙生草地含量仅为11.5 mg·kg-1,除沙生草地外,其他草地类型土壤速效氮含量最大值出现在10-20 cm土层;土壤全磷和速效磷含量在高寒草甸不同退化演替阶段及其各土层中差异不显著( P>0.05);高寒草甸退化演替过程中,土壤养分各因素间相互影响,其中,土壤全氮、全磷、速效氮和速效磷与土壤有机质之间呈显著正相关关系,土壤速效氮与全氮和土壤速效磷与全磷之间亦呈显著正相关关系,而土壤pH值与土壤各养分之间的相关性均不显著。
Soil nutrient is the basis for maintaining alpine grassland ecosystem health. The distribution characteristics of soil nutrient for swamp meadows, typical meadow, steppe meadow and sandy grassland were studied to provide theoretical basis for reasonable management of alpine grassland. The results demonstrated that (1)obvious difference was detected for soil organic matter and total nitrogen content between different type alpine grassland ecosystems, and swamp meadows>typical meadow>steppe meadow>sandy grassland. Moreover, soil organic matter and total nitrogen content showed a significant decrease trend with the increase of soil depth in all the studied alpine grassland ecosystems. (2) The soil available nitrogen content in steppe meadow was as high as 65.5 mg·kg-1, which was greater than those of the other three grassland ecosystems. In contrast, the soil available nitrogen content in sandy grassland was as low as 11.5 mg·kg-1, which was the least one. The soil available nitrogen content in typical meadow was greater than that in swamp meadow. Furthermore, the maximum value of soil available nitrogen among different alpine grassland ecosystems appeared in 10―20 cm soil layer except for the sandy grassland. (3) No significant difference for soil total phosphorus and available phosphorus were detected between different degradation periods of alpine grassland ecosystems as well as different soil layers of the same alpine grassland ecosystems. (4) Different soil nutrients could influence each other in the degradation process of alpine grassland ecosystem. A significant positive correlation was found between soil organic content and soil total total nitrogen, total phosphorus, available nitrogen and available phosphorus, as well as between soil available nitrogen and total nitrogen. In addition, a significant positive correlation was detected between soil available phosphorus and total phosphorus. However, no significant correlation could be found for soil pH and soil nutrients.
长江源区位于青海省南部, 面积达15.9万km2, 约占整个三江源区面积(36.3万km2)的43.8%, 是国家三江源自然保护区的重要组成部分[1]。高寒生态系统是该区最大的陆地生态系统, 其健康状况事关长江水源补给、区域生态安全和畜牧业生产。土壤养分供给是高寒草地植物健康生长的基础, 然而不同高寒草地类型间土壤养分供给能力存在一定的差异, 主要是不同类型高寒草地内植物组分存在差异, 其生长过程中对土壤养分的消耗不同。虽然土壤养分含量能够随着有机质分解而得到补充, 但不同类型高寒草地生境的分异, 决定了不同高寒草地类型间土壤养分的补给能力存异。查清不同类型高寒草地内土壤养分的分异性, 就能够因草地类型而采取合理管理和利用草地资源的策略。
土壤有机质是土壤养分的主要来源, 因此土壤有机质含量预示着土壤养分潜力的变化, 而青藏高原地区对高寒植物生长起重要作用的矿质元素主要是土壤氮和磷, 其中全氮和全磷反映了土壤养分的基础, 而速效氮和速效磷则反映了土壤供给植物养分的真实能力[2, 3, 4, 5]。长江源区高寒草地的类型主要包括沼泽化草甸、典型草甸、草原化草甸和沙生草地, 不同高寒草地植物组分随着土壤养分的变化而协同变化[6, 7, 8], 因此, 分析长江源区不同类型高寒草地间土壤有机质、土壤氮磷的变化特征, 对制定合理草地管理策略、维系草地持续利用具有重大的实践指导意义。
研究区位于长江源区腹地, 通天河直门达水文站以上长江干支流的集水区域, 范围介于90° 43'-96° 45' E, 32° 30-35° 35' N。研究区平均海拔超过4 000 m[4]。该地区地形地貌复杂多样, 生物资源丰富, 属于典型的高原大陆性气候, 气候基本特征表现为日温差大、年温差小、辐照时间长、辐射强度大, 冷暖两季较为漫长。年均气温在-1.7~5.5 ℃, 年均降水量在270~410 mm[9]。土壤类型以高山草甸土为主[10]。
2008年8月中旬, 沿青藏公路沿线设置样地, 选择不同的草地类型设置样地时, 每个各样的海拔、坡向、地表情况一致。其中沼泽化草甸、典型草甸、草原化草甸和沙生草地分别设置样地4个、10个、12个和9个, 用GPS测定样地的经纬度和海拔。沼泽化草甸的建群种为藏嵩草(Kobresia tibetica), 由耐寒湿生和湿中生多年地面芽和地下芽植物组成, 以莎草科、禾本科和毛茛科植物为主, 总盖度90%以上。典型草甸建群种为矮嵩草(K. humilis)和线叶嵩草(K. capillifolia)等中生植物, 总盖度50%~70%。草原化草甸群落的优势植物为小嵩草(K. pygmaea), 湿生植物消失, 草原植物出现, 总盖度45%~55%。沙生草地中生草甸植物消失, 主要以耐旱植物为主, 紫花针茅(Stipa purpurea)和青藏苔草(Carex moorcroftii)占据绝对优势, 总盖度低于40%[11]。
在每个样地内, 随机设置1 m× 1 m的样方5个。收获地上植被后, 采用内径5 cm 土钻分3层(0-10, 10-20和20-40 cm)取土, 每个样方按“ V” 字型钻取5钻, 每个样地的每个土层共25钻, 然后将同一土层土样混合为一个土样, 形成75个土样, 编号后用透气的布袋带回实验室, 过2 mm土筛, 并仔细剔除细根等杂物后, 进行土壤理化性质测定。其中0-40 cm土层土壤参数依据郭正刚等[11]采用平均值, 便于和其他地区相关土壤参数进行比较。土壤有机碳用磷酸浴— 重铬酸钾容量法, 全氮和速效氮采用凯氏法和康维皿法, 全磷采用钼锑抗比色法, 速效磷采用碳酸氢钠浸提-钼锑抗比色法, 土壤pH值用酸度计测定[12]。
用Excel 2013和SAS 9.3软件进行数据的统计分析, 数据用ANOVA方差分析, 差异显著性运用LSD检验法进行多重比较。利用Origin 8.0进行绘图。
不同高寒草地类型间土壤有机质含量存在显著差异(P< 0.05)(图1), 0-10、10-20和20-40 cm土层土壤中有机质含量均表现为沼泽化草甸> 典型草甸> 草原化草甸> 沙生草地, 且随土层深度加深, 典型草甸、草原化草甸与沙生草甸的土壤有机质含量均呈现降低的变化态势, 而沼泽化草甸却出现一定的分异, 随土层加深其土壤有机质含量先增加后降低, 在10-20 cm土层土壤中达到最大。
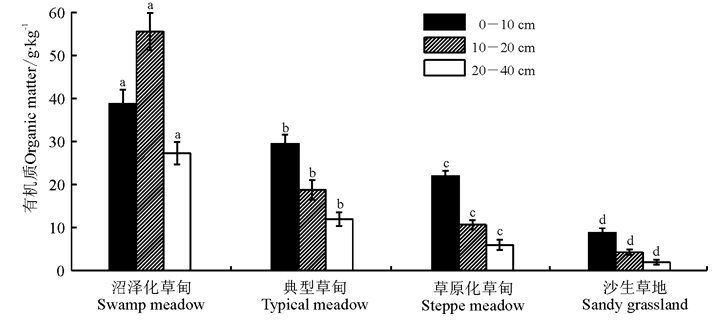 | 图1 不同高寒草地类型土壤有机质含量注:不同小写字母表示相同土层不同草地类型间差异显著(P< 0.05)。下同。Fig.1 The soil organic matter content of different alpine grasslandNote: Different lower case letters for the same soil layer indicate significant difference among different alpine grasslands at 0.05 level. The same below. |
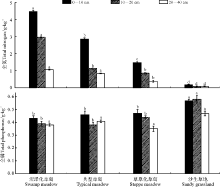
土壤全氮和全磷含量在不同高寒草地类型间均具有明显的差异(图 2), 具体表现为各土层全氮含量均呈沼泽化草甸> 典型草甸> 草原化草甸> 沙生草地, 且随土壤土层加深, 各高寒草地类型的土壤全氮含量均呈降低的变化趋势。土壤全磷含量在0-10 cm土层表现为沙生草地> 草原化草甸> 典型草甸> 沼泽化草甸, 10-20 cm土层表现为沙生草地> 草原化草甸> 沼泽化草甸> 典型草甸, 20-40 cm土层土壤全磷含量表现为沙生草地> 典型草甸> 沼泽化草甸> 草原化草甸, 但0-40 cm土层总体表现为沙生草地(0.54 g· kg-1)> 草原化草甸(0.44 g· kg-1)> 典型草甸(0.42 g· kg-1)> 沼泽化草甸(0.40 g· kg-1); 随土壤深度加深, 在沼泽化草甸和草原化草甸中, 土壤全磷含量逐渐降低, 而在典型草甸中土壤全磷含量表现为先降低后增加的变化趋势, 在10-20 cm土层中最少; 在沙生草地中随土壤深度加深土壤全磷含量呈先增加后降低的变化特征, 20-40 cm土层中最少。
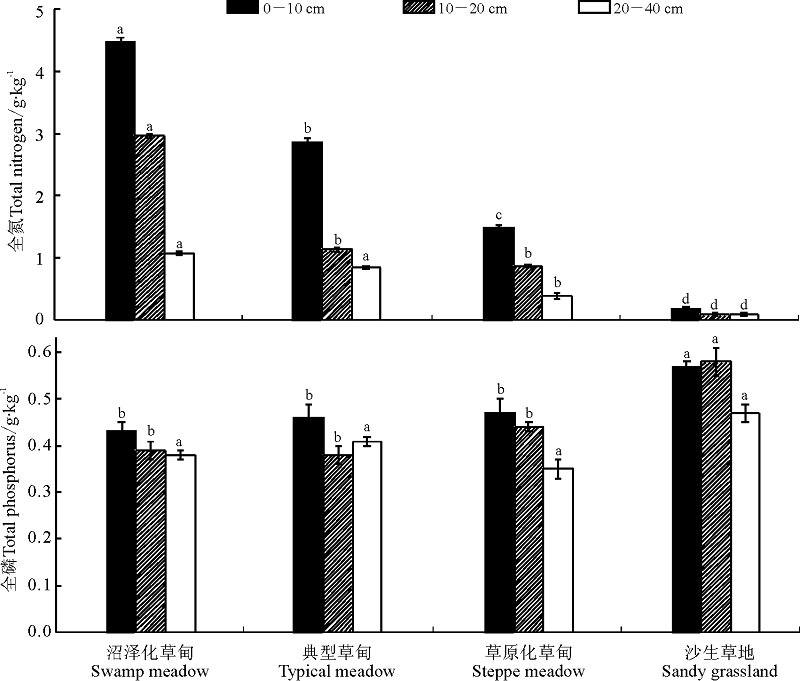 | 图2 高寒草甸不同退化演替序列土壤全氮和全磷含量变化Fig. 2 Changes of total N and total P in the different degradation succession stages in the alpine meadow |
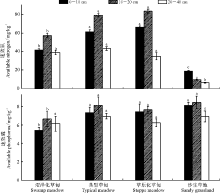
结果表明, 土壤速效氮和速效磷含量在不同高寒草地类型间明显不同(图 3), 0-10、10-20 cm土层和0-40 cm土层中, 土壤速效氮含量表现为草原化草甸> 典型草甸> 沼泽化草甸> 沙生草地, 而在20-40 cm土层土壤中, 速效氮含量却呈现典型草甸> 沼泽化草甸> 草原化草甸> 沙生草地的变化特征, 随土层深度的增加, 除沙生草地土壤速效氮表现为降低态势外, 其余各高寒草甸类型的土壤速效氮均随土层深度的增加呈现先增加后降低的变化趋势。4种草地类型各土层土壤速效磷含量的变化特征具体表现为:0-10 cm土层中, 沙生草地> 草原化草甸> 典型草甸> 沼泽化草甸; 10-20 cm土层中典型草甸> 草原化草甸> 沼泽化草甸> 沙生草地; 20-40 cm中, 沙生草地> 典型草甸> 沼泽化草甸> 草原化草甸; 0-40 cm土层总体上表现为, 沙生草地(7.8 mg· kg-1)> 典型草甸(7.4 mg· kg-1)> 草原化草甸(7.1 mg· kg-1)> 沼泽化草甸最低(6.0 mg· kg-1)。
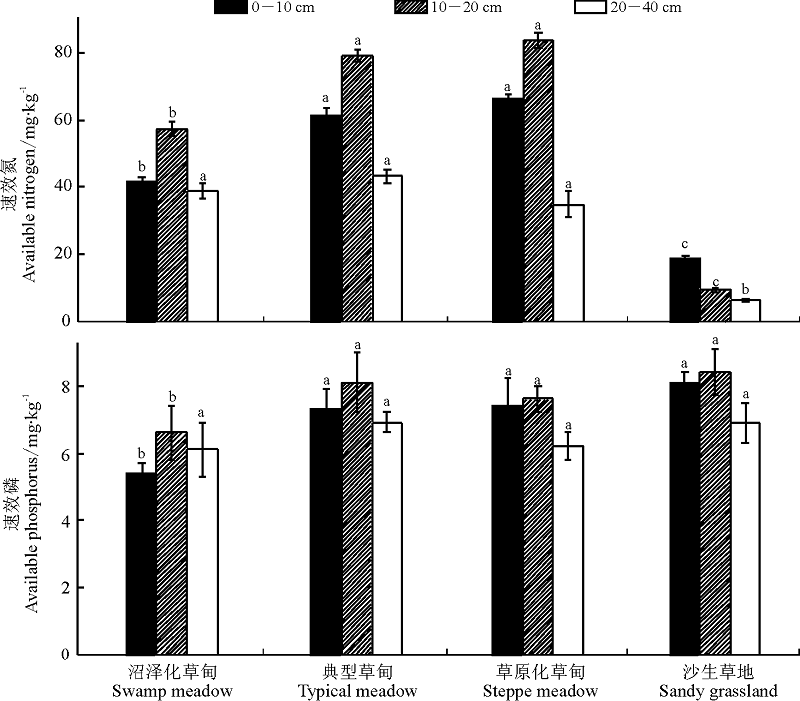 | 图3 土壤速效氮和速效磷含量的变化Fig.3 Changes of soil available nutrient content matter content in the different degradation succession stages in the alpine meadow |
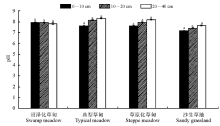
土壤pH值是控制土壤养分循环的重要因子, 4种草地类型各土层土壤pH值(图4)为:0-10 cm, 沼泽化草甸> 草原化草甸> 典型草甸> 沙生草地; 10-20 cm, 典型草甸> 沼泽化草甸> 草原化草甸> 沙生草地; 20-40 cm, 典型草甸> 草原化草甸> 沼泽化草甸> 沙生草地。0-40 cm, 典型草甸(8.03)> 草原化草甸(7.92)> 沼泽化草甸(7.91)> 沙生草地(7.41)。
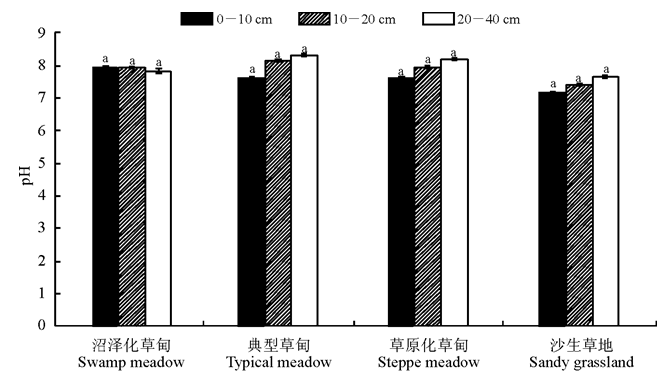 | 图4 高寒草甸不同退化演替序列土壤pH值的变化Fig.4 Changes of soil pH value in the different degradation succession stages in the alpine meadow |
4种草地类型土壤有机质和全氮(r=0.978)、全磷(r=0.813)、速效氮(r=0.732)以及速效磷(r=0.738)含量均呈显著正相关(表1), 土壤速效氮与全氮(r=0.713)、土壤速效磷与全磷(r=0.987)之间亦呈显著正相关关系, 这说明有机质是决定草地土壤养分的关键因子, 土壤速效养分含量受土壤全量养分的调控。
| 表1 土壤养分之间的相关关系 Table 1 Correlation relationship between the soil nutrient content |
土壤有机质不仅是土壤主要的养分指标, 也是形成土壤结构的重要因素, 能够直接影响土壤肥力、持水力以及抗侵蚀的能力[13], 其主要来源于植物残体的凋落分解, 不同草地类型会导致土壤有机质含量的不同[14]。研究结果表明, 土壤有机质含量呈沼泽化草甸> 典型草甸> 草原化草甸> 沙生草地的变化特征, 主要原因是不同高寒草地类型生产力不同, 植物残体有差异, 有机质输入出现一定的分异, 此外不同高寒草地类型分解有机质的速率也不同。王根绪等[3] 研究发现长江源区4种草地类型植物生物量差异显著, 植物归还量依次是沼泽化草甸> 典型草甸> 草原化草甸> 沙生草地。另外, 在沼泽化草甸中, 多年冻土层保存完好, 它的存在能有效阻止土壤水分和地表水的下渗, 致使植被根系层土壤的水分含量较高, 增加了土壤的保水保肥能力, 加之沼泽化草甸土壤温度较替他演替阶段低, 微生物活动受限, 有机质分解受到抑制, 有机质在该类型草地中大量聚集。然而在高寒草地类型从典型草甸到草原化草甸再到沙生草地的变化过程中, 冻土活动层逐渐加深, 多年冻土上限逐渐下降, 使得植被根系层土壤的水分含量依次下降, 土壤保水保肥能力逐渐降低[15]。同时, 随草地类型的变化(典型草甸→ 草原化草甸→ 沙生草地), 土壤温度逐渐升高, 微生物活性增加, 分解有机质的速率增加, 使土壤有机质含量不断下降[16]。因此, 在高寒草地类型变化过程中, 土壤有机质含量表现为沼泽化草甸> 典型草甸> 草原化草甸> 沙生草地的特征。研究发现, 典型草甸、草原化草甸和沙生草地土壤有机质的含量有着明显的垂直分布规律:各土壤层有机质含量随土壤深度增加呈显著降低趋势。而沼泽化草甸土壤有机质含量的最大值却出现中层(10-20 cm), 该结果和王彦龙等[17]的研究结果相似, 沼泽化草甸是大气CH4的重要来源, 在水淹等缺氧环境下, 土壤上层储存的有机碳会在微生物的作用下发酵产生CH4并释放到大气中, 造成有机碳损失[18]。
土壤氮素主要以有机氮的形式存在于表层土壤中, 土壤有机碳含量能够显著影响土壤氮素矿化潜力[19], 当可利用性碳源相对充足时, 氮的固定潜能高, 净氮矿化降低, 增加全氮含量; 相反, 当可利用性碳相对缺乏时, 氮的固定潜能低, 净氮矿化升高, 加速全氮分解[20], 受土壤有机质含量的驱动, 土壤全氮含量的变化趋同于土壤有机碳含量的变化, 而土壤有机碳来源于土壤有机质, 且研究发现有机质是决定草地土壤养分的关键因子, 因此4种草地类型间土壤全氮含量的变化特征与有机质含量变化特征相一致。
土壤中的磷多是来自母岩矿物[19], 而研究区土壤母岩矿物基本相同。因此各高寒草地类型土壤全磷含量与草地生产力密切相关。沼泽化草甸的生物量最高, 沙生草地生物量最低, 而典型草甸和草原化草甸生产力介于二者之间, 因此沼泽化草甸对磷素的需求较其草地类型高, 所以不同高寒草甸类型间土壤全磷含量变化趋势表现为沙生草地> 草原化草甸> 典型草甸> 沼泽化草甸。然而不同高寒草地类型土壤全磷含量垂直分布不明显, 各土层全磷在含量差异不明显, 说明草地类型对多年冻土区草地土壤全磷的影响相对较小。
土壤中的速效养分能够直接被植物利用, 对植物的生长具有迅速而直接的影响, 是高寒草地植物生长过程的主要限制因子[20, 21]。土壤中的速效氮含量主要土壤氮矿化速率的影响。高寒草甸类型从沼泽化草甸→ 典型草甸→ 草原化草甸→ 沙生草地转变的过程中, 草甸下面冻土和地表给水逐渐减少, 土壤表层水含量逐渐下降, 土壤蒸散量逐渐减少, 从而使由于蒸散损失的热量减少, 土壤温度逐渐升高。而土壤温度的升高促进了土壤微生物的活动, 加速了土壤氮素的矿化作用。因此较低的土壤温度使沼泽化草甸土壤速效氮含量低的主要原因。然而由于沙生草地的植被覆盖度较低, 土壤中植物根系减少, 土粒松散, 土壤氮素大量流失[20, 21], 因此, 典型草甸和草原化草甸土壤速效氮含量显著高于沼泽化草甸, 而沙生草地速效氮含量显著低于其他类型草地。和沙生草地不同, 沼泽化草甸、典型草甸和草原化草甸土壤速效氮含量的最大值出现在土壤中层(10-20 cm), 主要原因是一方面这3类草地的植物主要是浅根系植物[10], 在其生长过程中对上层土壤(0-10 cm)氮素的吸收能力较强, 另一方面高寒草地生态系统土壤速效氮组分主要以氨态氮为主, 由于土壤上层易受外界环境的影响, 氨的挥发损失最严重, 使其中相当数量的氮素被大大地浪费掉[22]。
土壤速效磷在沙生草地最高, 典型草甸和草原化草甸次之, 沼泽化草甸最低, 这与植物生物量从沼泽化草甸、典型草甸和草原化草甸, 到沙生草地依次降低有关, 因为植物种类的减少降低了对土壤磷素的摄取。土壤速效磷含量受土壤全磷含量的约束, 因此, 不同高寒草地土壤速效磷含量和土壤全磷的变化特征趋同。
长江源区不同高寒草地类型的土壤有机质和氮磷含量均存在明显差异, 其中土壤有机质和全氮含量表现为沼泽化草甸> 典型草甸> 草原化草甸> 沙生草地, 速效氮含量却表现为草原化草甸> 典型草甸> 沼泽化草甸> 沙生草地, 但土壤磷含量在不同高寒草地类型间差异不明显。因此高寒草地管理时应该充分考虑有机质输入量和氮素的有效性, 而磷元素的作用不是非常明显。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|