
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
亚高寒草甸物种内稳性与生物量变化模式
[张仁懿1  , 史小明
, 史小明1 , 李文金1 , 郭睿2 , 王刚1 ]
, 史小明|
|


植物的内稳性与其在群落中的生态策略和适应性密切相关,然而从物种或功能群水平探讨内稳性与其竞争力关系的研究非常有限。本研究以青藏高原亚高寒草甸为例,通过N添加、P添加及N-P共添加的方式,选取4个功能群的10个物种,探讨养分添加下生物量变化规律及物种内稳性差异。结果显示,N添加及N-P共添加促进生物量的积累,而P添加抑制群落生物量的积累,说明该地区主要限制类型为N限制;N添加、P添加及N-P共添加均促进禾本科植物生物量的积累,而降低豆科及杂类草植物的生物量,其中仅N添加增加莎草科植物的生物量;P添加下,禾本科植物P内稳性强于其他物种,P内稳性可能在禾本科植物的竞争力上起到重要作用;其他处理及N、N:P比未表现出一般性规律。因此,养分添加条件下,内稳性差异在高寒草甸物种竞争中可能并非主导因素。
Plant stoichiometric homoeostasis is correlated with dominance and stability in natural community. However, there is considerable limited work dealing with this relationship at species-level or function group-level. Study was made on the response of 10 dominant speicies from four function groups to N, P addition and N-P co-addition on a sub-alpine meadow in Tibetan Plateau. Results showed N and N-P addition promoted community biomass while P addition inhibited, indicating the N-limited type for grassland. Graminoid biomass increased, while legume and forb biomass decreased after either addition, and sedge biomass increased after N addition. The strong leaf P homeostasis of graminoid species may play a role in competition in the community. However, a regular rule on the leaf N homeostasis needs to be found out, and further study should be conducted to test the role of species homeostasis in the construction of community.
植物具有调整自身N:P 化学计量特征, 以适应环境养分供应条件变化的能力。生物体内营养物质的浓度过高或者过低, 都将影响其生存与生长:浓度过低将使生物体停止生长并启动衰老过程, 而过高则会对植物机体产生毒害作用[1]。植物经过长期的适应与进化, 发展了维持体内养分含量相对稳定的能力。即使外界环境不断变化, 其组成也不会发生很大变化, 这种有机体与其环境保持一种相对稳定状态的现象就被称为“ 内稳态(Homeostasis)” [2]。内稳性是生态化学计量学的核心概念, 这个指标反映了生物对环境变化的生理和生化的适应, 其强弱与物种的生态策略和适应性有关[3]。例如, Yu等[4]针对内蒙古草原1 200 km 横断范围、为期27 年的研究表明, 生态化学计量内稳性高的物种在群落中具有较高的优势度与稳定性, 因此内稳性成为衡量物种竞争力的一个重要指标。
养分添加破坏了原有的养分供需平衡, 进而影响种间关系的作用, 引起物种组成和群落结构的变化[4]。高寒、亚高寒草甸自然群落的养分添加试验表明, 不同物种、不同功能群植物对养分添加的响应不同[5], 这与他们之间N:P化学计量特征尤其是内稳性存在的差异密切相关[1]。关于生态化学计量内稳性的生态学和进化学意义, 已得到生态学学者们的重视, 但研究成果十分有限[3]。本研究设置了3个N-P养分添加组合, 4个水平的添加量的处理, 选取莎草科、禾本科、豆科及杂类草4个功能群的10个代表物种, 观察物种化学计量特征及其生物量对养分添加的响应, 讨论物种化学计量内稳性在群落构建中的意义。
研究地点位于兰州大学高寒草地生态系统定位研究站合作站(34° 56' N, 102° 52' E), 海拔2 900 m。站点属青藏高原东北部, 为寒温湿润的高原气候。根据合作气象站的资料显示, 该地年平均气温2.4 ℃, 年平均降水量为530 mm, 年平均蒸发量1 200 mm, 植被类型属于多年生草本植物占优势的亚高寒草甸。
2008年秋季, 选取试验站禁牧区内面积53 m× 35 m的样地, 坡度约为3° 。将样地划分为54个5 m× 5 m的区块, 相邻区块间隔1 m避免边缘效应及作为过道(图1)。选取其中50个区块进行养分添加处理, 采用随机区组设计进行N添加、P添加、N-P共添加; 养分添加量为0、5、10、15 g· m-2。共10个处理, 每个处理设计5个重复(图1)。添加的养分均为速效肥, 其中N为尿素[CO(NH2)2 , N%=46%], P为磷酸二氢钠(NaH2PO4, P%=26%)。2008年及后续2009―2011年养分添加处理时间均为8月中旬。
 | 图1 样地布置图Fig.1 Distribution map of sampling plots |
2012年8月中旬生物量及物种样品的采集。各区块选择两个0.5 m× 0.5 m的样方, 分功能群(莎草科、禾本科、豆科及杂类草)齐地剪下生物量, 70 ℃烘干至恒重以计算生物量。各区块的生物量样方及长期观测样方外, 每个区块分别采集10个共有物种的叶片约5 g, 70 ℃烘干至恒重待分析。所选物种为各处理的共有种, 包括莎草科的高山嵩草(Kobresia pygmaea)、矮藨草(Scirpus pumilus), 禾本科的垂穗披碱草(Elymus nutans)、草地早熟禾(Poa pratensis), 杂类草的藏大戟(Euphorbia tibetica)、甘青老鹳草(Geranium pylzowianum)、兰石草(Lancea tibetica)、川甘蒲公英(Taraxacum lugubre)以及豆科的花苜蓿(Trigonenica ruthenica)、甘肃棘豆(Oxytropis kansuensis)。
植物样品经研磨后过0.180 mm筛, 采用H2SO4-H2O2消煮法消煮[6]。植物样品的N、P含量用仪器SmartChem 200(AMS-Westco)以比色法的原理分析测定。
使用线性回归来分析养分添加量与各生物量之间的关系, 使用Origin 8.0分析和作图; 采用一般线性模型(GLM)分析养分添加类型对地上生物量的交互作用; 物种内稳性通过养分添加量梯度下物种N-P化学计量特征的斜率衡量, 斜率为零时为严格的内稳态, 斜率越大内稳性越弱[7], 其中斜率计算用SPSS 13.0分析, 而作图均在Origin 8.0完成。
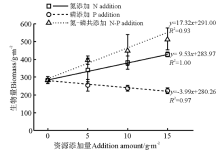
N添加、P添加或N-P共添加处理下, 群落水平的地上生物量均与养分添加量显著相关。其中, N-P共添加(P< 0.05)对群落生物量的促进作用强于N添加(P< 0.001); 而P添加(P< 0.05)抑制地上生物量的积累(图2)。GLM分析结果显示, N、P养分添加存在显著的交互作用(P< 0.05)。
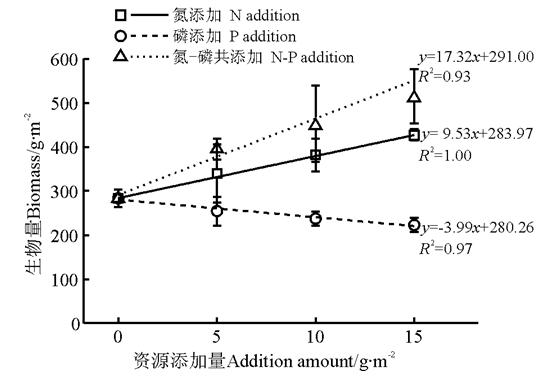 | 图2 养分添加量与地上生物量的回归关系Fig.2 Relationships between addition amount and aboveground biomass |
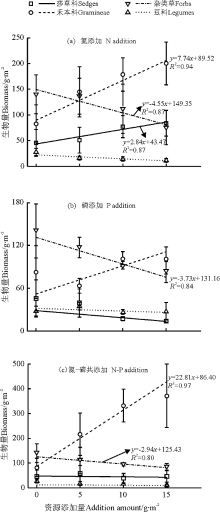
功能群水平, N添加促进莎草科生物量, 而P添加及N-P共添加抑制其生物量的积累(斜率分别为2.84、-1.00和-0.31, P> 0.05); 禾本科生物量与养分添加量呈正相关, 养分添加的促进作用顺序为N-P共添加> N添加> P添加, 斜率分别为22.81(P< 0.05)、7.74(P< 0.05)和3.94(P> 0.05); 杂类草生物量与养分添加量呈显著负相关, 养分添加的抑制作用顺序为N添加> P添加> N-P共添加, 斜率分别为-4.55(P< 0.05)、-3.73(P< 0.05)和-2.94(P< 0.05); 养分添加抑制豆科生物量, 抑制作用无显著差异, 表现为N添加> N-P添加> P添加, 斜率分别为-0.77、-0.38和-0.17(图3)。
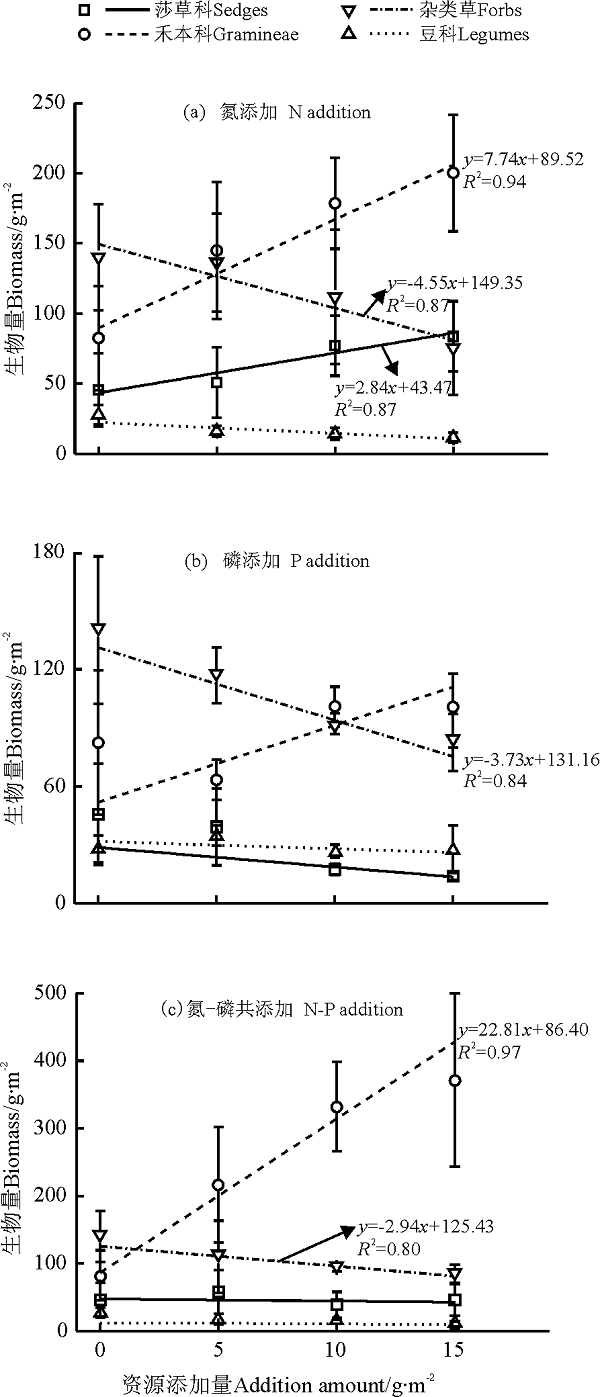 | 图3 养分添加量与各功能区地上生物量的回归关系Fig.3 Relationships between P addition, N addition and N-P co-addition amount and aboveground biomass of function groups |
物种水平的生物量变化模式与功能群的生物量变化模式一致。莎草科的高山嵩草及矮藨草生物量随N的添加量增加而上升, 而随P及N-P共添加的添加量增加而降低; 在N添加、P添加以及N-P共添加下, 禾本科植物(垂穗披碱草和草地早熟禾)生物量与养分添加量正相关; 除了P添加下的藏大戟和甘肃棘豆、N添加下的藏大戟和甘青老鹳草, 各养分添加组合下, 豆科物种和杂类草物种生物量均随着养分添加量的增加而下降(表1)。
| 表1 样地10个物种叶片N、P、N:P以及生物量与养分添加量线性回归的斜率值 Table 1 Slopes of the linear regression between species traits (N, P, N:P ratio and biomass) and addition amount for 10 species |
对于所有选择物种, 随着P添加量的增加, 叶片P含量上升, N:P比值下降; 随着N添加量的增加, 叶片N含量及N:P比值也上升; 除了甘肃棘豆叶片P含量接近0外(斜率为-0.01), N-P共添加表现出与P添加一致的影响, 即随添加量的增加, 除甘肃棘豆外, 其余物种叶片P上升, N:P比下降, 然而各物种P含量、N:P比值随养分添加的变化斜率较单一P添加的斜率小。
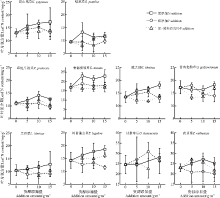
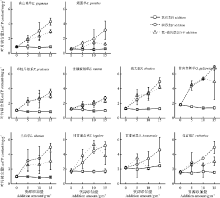
单一P养分添加下, 禾本科植物垂穗披碱草和草地早熟禾的P内稳性在所选物种中最强; 然而, 在N添加及N-P共添加处理下, 物种间及功能群间的内稳性差异在本研究中难以找到特定的规律, 物种叶片N、P以及N:P的内稳性与其生物量变化之间无显著的相关关系(表1, 图4、5、6)。
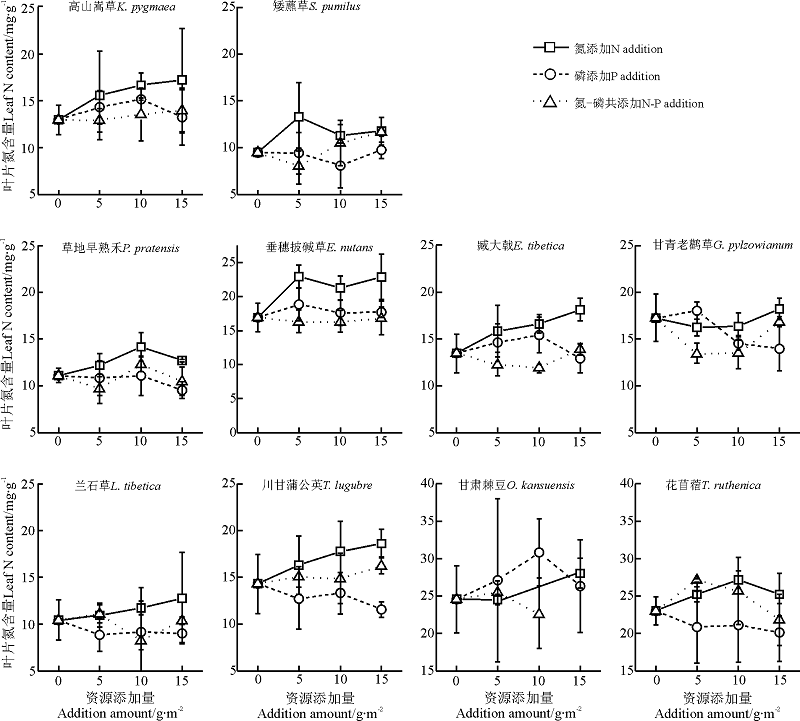 | 图4 各养分添加组合及添加量下10物种的叶片N含量Fig.4 Leaf N contents of 10 selected species after P, N addition and N-P co-addition with different addition ammounts |
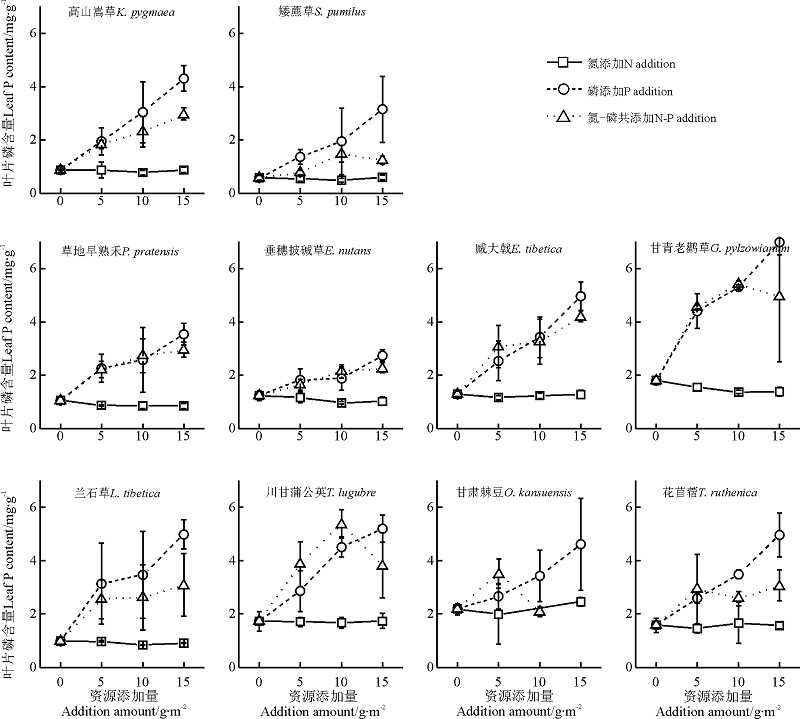 | 图5 各养分添加组合及添加量下10物种的叶片P含量Fig.5 Leaf P contents of 10 selected species after P, N addition and N-P co-addition with different addition amounts |
 | 图6 各养分添加组合及添加量下10物种的叶片N:P比值Fig.6 Leaf N:P ratios of 10 selected species after P, N addition and N-P co-addition with different addition ammounts |
青藏高原亚高寒草甸养分添加结果显示, N添加显著促进地上生物量的积累, 而P添加显著抑制地上生物量的积累(图2), 表明该地区自然植被以N限制为主要的养分限制类型, 这与其他地区高山地区限制因子的研究结果一致[8]。土壤中植物可吸收的N素主要来源于N的矿化, 而此过程为生物代谢过程, 其对温度的敏感度远高于土壤P等养分的释放[9], 因此, 亚高寒草甸所处的低温环境使该地区植被受N供应的限制大于其他元素的影响[10, 11]。P添加对生物量的抑制作用, 表明群落中多数物种对P素的吸收已达到饱和状态, 甚至受到了机体内P素过度积累的毒害。
亚高寒草甸N-P养分的共添加存在交互作用(图2), 说明N素养分的添加, 能够减弱或消除P素过量积累的毒害作用, 这与群落中各组分限制因子及养分利用方式的多样化密切相关[8, 11]。例如, 豆科植物常常与固氮菌形成互惠共生体, 这个共生体使多数豆科植物具有额外的N素来源, 易受P素缺乏的影响[12, 13]; 而非豆科植物一般具有较高的N素利用效率, 相对于豆科植物, 更易受N素供应的限制[1, 11]。N-P养分的共添加, 在一定程度上缓解了这两个类群植物的养分限制, 从而获得了较高的生物量[14, 15]。
养分的添加改变了原有自然植被的养分供应-消耗平衡, 群落中不同物种、不同功能群植物对此作出的响应存在差异, 从而改变原有群落的物种组成及群落结构[14, 16]。对亚高寒草甸植被长期养分添加的结果显示, 单一的N、P添加或是N-P养分的共添加, 禾本科植物都能做出积极的响应, 增强其在群落中的主导作用。这与其他高寒、亚高寒草甸地区的研究一致[14, 17]。养分添加促进禾本科植物的生长, 提高禾本科植物在群落中的优势度, 这在许多植被类型中已得到了有效验证[9, 18, 19]。
内稳性是生态化学计量学存在的理论基础, 其强弱与物种的生态策略和适应性有关[3]。在内蒙古草原上的研究表明, 生态化学计量内稳性高的物种在群落中具有较高的优势度与稳定性, 是物种竞争力的一个重要指标; 而群落的内稳性则是生态系统功能健康、群落结构稳定的体现[4]。以外界养分含量为横坐标, 机体内养分含量为纵坐标, 其斜率与物种内稳性互为倒数关系。该斜率等于0则为绝对的内稳态, 内稳性无穷大, 接近0则内稳性强, 反之斜率越大则内稳性越弱[7]。P添加下所选物种的P斜率及N添加下物种N斜率均大于0, 说明所选物种均不具有绝对的内稳性, 机体内养分含量随养分供应变化而发生调整。
N-P共添加对所选物种生态化学计量学特征的影响主要表现为叶片P含量的变化, 物种水平的内稳性与单一P添加相近, 但养分添加量与叶片P含量回归的斜率小于P添加。这进一步说明该地区的物种尤其是非豆科植物的P供应处于饱和状态, 植被的主要限制因子为N限制[8, 15]。P富足的植被添加P养分后, N的共添加缓减了P过量对植物的毒害作用[20]。对于亚高寒草甸P富足的土壤, 植物的P内稳性在提高物种适合度的过程中显得尤为重要。研究结果显示, 对N、P养分添加最能作出积极响应的两个种为禾本科的垂穗披碱草和草地早熟禾, 他们具有最低的P斜率即最强的P内稳性。然而, N养分添加下N的内稳性在不同功能群的物种间没有找到一般性规律, 难以用N内稳性差异解释物种、功能群之间生物量变化模式的差异。
禾本科植物在单一N、P养分添加及N-P共添加下均表现出较高的适应性。P内稳性可能在禾本科植物的竞争力上起到重要作用, 然而, 本研究并未发现物种N内稳性与生物量间的相关性。养分添加下禾本科植物的主导作用, 内稳性仅仅是众多影响因素的一小方面。相对其他草本植物, 禾草植物普遍具有发达的根系, 能够快速而有效地吸收所添加的养分[21, 22]。因此, 在N限制的亚高寒草甸自然N梯度上, 禾本科植物在低N土壤上更具竞争力[15]。此外, 禾本科植物的克隆特性及植株高度快速增加, 也是他在养分添加后占据主导地位的重要因素[23]。
生态化学计量内稳性是生态系统结构、功能和稳定性维持的重要机理, 但养分添加可能会改变生态化学计量内稳性与生态系统特性的关系[3]。因此, 为了更准确地验证内稳性与物种适应度之间的关系, 后续研究应减小人为干扰, 以自然养分条件为梯度, 以及选择更多物种以利用有效的统计方法。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|