
荒漠草原4种优势植物叶片化学计量内稳性特征
探讨植物生态化学计量内稳性特征对了解草原植物的生长适应策略具有重要意义。以内蒙古鄂克托前旗荒漠草原牛枝子(Lespedeza potaninii)、苦豆子(Sophora alopecuroides)、短花针茅(Stipa breviflora)和猪毛蒿(Artemisia scoparia) 4种优势植物为研究对象,研究植物叶片氮、磷内稳性特征及其影响因素。结果表明:苦豆子叶片N、N ꞉ P含量显著高于其他3种优势植物(P < 0.05),猪毛蒿叶片P含量显著高于其他3种优势植物;N元素内稳性指数(HN)以牛枝子最高,属于稳态型;P元素内稳性指数(HP)以猪毛蒿最高,属于稳态型;N ꞉ P内稳性指数(HN ꞉ P)以牛枝子最高,属于弱稳态型。冗余分析结果显示,土壤全碳和全氮含量对化学计量内稳性变异解释率分别为56.9%和9.9%,均达到显著水平(P < 0.05),但其他土壤理化因子对其无显著影响(P > 0.05)。综上所述,在氮含量较低的荒漠草原,多年生植物N内稳性和N ꞉ P内稳性较高,采用保守养分策略,其中N内稳性较高的灌木牛枝子适应性更强;一年生植物P内稳性较高,N ꞉ P内稳性比较低,采用奢侈分配策略;影响化学计量内稳性主要因子为土壤全碳和全氮含量。
English
-
作为陆地生态系统的基本组成部分,植被充分接触大气、水和土壤,并通过调节水循环、碳循环和能量循环来推动生态系统服务[1]。对于制订可持续生态系统管理策略来说,遥测植被的时空动态变化至关重要[2]。植被覆盖度(fractional vegetation cover, FVC)是指地面植被垂直投射面积与区域总面积的比例[3],可以了解地表植被状况以及区域环境所存在的问题[4]。20世纪以来,随着人口增加和城镇化建设的推动,耕地、建设用地扩张、滥砍滥伐滥牧等人类活动对植被的影响日益显著[5],工业、农业生产建设从自然界中获取大量原材料,导致全球森林与草原植被变化呈现下降趋势,生态系统的功能和结构遭到严重损害引发的生态危机日益加重[6]。因此及时准确探测植被覆盖度的时空变化,对自然资源可持续利用、经济可持续发展和生态恢复都具有重要应用价值[7],同时探讨不同驱动因子对地表植被时空变化的影响已成为植被系统方面的重点[8]。
随着生态文明建设进程的不断推进,植被变化的驱动力分析已成为国内外学者关注的热点之一,近年来已有很多结论,如张一然等[9]探讨若尔盖湿地植被覆盖时空变化和气候因子的影响机理,结果表明植被覆盖度变化与气温、降水均呈正相关关系。吉珍霞等[10]基于长时间序列的MODIS数据解析了青藏高原草地物候变化并了解其中的驱动机制,结果表明影响草地生长季长度的主要作用因素是风速和连续5 d最大降雨。以往研究多采用相关系数分析法[11-12]来定性分析自然因素和人为因素对植被覆盖度时空变化的影响,但很少定量分析驱动因子间的交互作用对植被覆盖度空间分层异质性的影响,且研究影响因子之间的协同作用较少[13]。地理探测器是一种新的分析因子与驱动力的统计方法,用于度量空间分异性,定量不同因子及其交互作用[14]。近年来,学者们运用该模型研究植被覆盖变化及驱动机制,目前主要分为3类:一是辨别主要的影响因素,如Yuan等[15]研究发现降水量是影响黑河流域归一化植被指数(normalized difference vegetation index, NDVI)变化的主导因素。二是研究驱动因子之间的耦合作用,如Liu等[16]定量分析潜在驱动因素对青藏高原东北部典型高寒地区的植被动态的影响,分析得出温度与土壤类型的协同作用对植被变化的影响最强。三是分析各影响因子对植被生长的适宜类型或范围,如Wang等[17]通过地理探测器模型研究发现促进植被生长的各自然因子最佳范围以及该条件下FVC均值具有显著的差异。
开都-孔雀河流域是干旱区内陆河塔里木河“四源一干”的重要源流之一[18],也是“丝绸之路经济带”建设的关键区。其气候干旱,生态环境脆弱,属于干旱区敏感地带。近年来人口增加[19],耕地、城乡工矿居民用地面积大范围扩张[20-21],焉耆盆地田间灌水量增加[22],导致孔雀河下游水资源短缺[23],植被退化严重[24],危及到流域的生态系统安全,并导致一系列严重的生态环境问题。本研究把开都-孔雀河流域作为研究靶区,通过Sen + Mann-Kendall趋势分析法对2000-2020年开都-孔雀河流域的FVC时空变化特征进行分析,同时定量分析各种自然和人为因子对植被时空变化的作用机理,为该流域的植被和生态恢复提供科学依据。
1. 数据与方法
1.1 研究区概况
开都-孔雀河流域位于新疆巴音郭楞蒙古自治州(简称巴州),地理坐标为82°55′~90°25′ E、39°35′~43°30′ N,研究区总面积为115 403.88 km2,行政区包括库尔勒市、尉犁县、轮台县、和硕县、博湖县、焉耆回族自治县、和静县、若羌县8个县(市)(图1)。至2020年,研究区总人口数达113.67 万人,GDP达9 453 671 万元[25]。该流域由开都河流域、孔雀河流域组成,开都河是自北向南注入博斯腾湖,出湖口后流入孔雀河。开都-孔雀河流域处于塔里木盆地东北部,西接塔克拉玛干沙漠,东靠库鲁克沙漠,属于典型的温带大陆性干旱气候,气候特征表现为昼夜温差大、降水量稀少、蒸发剧烈。
1.2 数据来源
1)遥感数据:NDVI数据来源于美国NASA EOS/MODIS (https://Ipdaac.usgs.gov/api)发布的MOD13Q1数据产品,分辨率为250 m × 250 m,间隔为16 d,研究区覆盖共有两景影像,分别为h24v04、h24v05,每年共计23个时相数据。该数据已经过水、云、气溶胶等去除处理,通过MODIS Reprojection Tool工具对数据进行一系列处理,获得2000-2020年NDVI数据。
2)气象数据:来源于中国气象科学数据共享服务网(http://cdc.cma.gov.cn),提取研究区内的标准气象站点的月平均数据和月降水数据。根据各气象站点的经纬度信息,对气象数据进行反距离权重插值,获得2020年的年均气温和年降水量。
3)地形数据:数字高程模型(digital elevation model, DEM)来源于地理空间数据云(http://www.gscloud.cn),从该数据中提取高程和坡向。
4)土地利用数据:选自中国科学院资源科学与数据中心(https://www.resdc.cn)公布的2020年30 m空间分辨率的土地利用数据。
5)社会经济数据:均来源于2020年的《新疆统计年鉴》 [25],提取各县(市)的数据,在Arcgis中进行空间关联并生成矢量数据。
1.3 研究方法
1.3.1 植被覆盖度
本研究选择了被广泛用于从NDVI计算FVC的像元二分模型,计算公式如下[26]:
$ FVC = \dfrac{{NDVI - NDV{I_{{{{\rm{soil}}}}}}}}{{NDV{I_{{{{\rm{veg}}}}}}-{{ NDV}}{{{I}}_{{{{\rm{soil}}}}}}}} 。 $

(1) 式中:NDVIveg表示纯植被区域的NDVI,NDVIsoil表示纯裸土区域的NDVI。根据影像提取结果和实际情况,本研究选取累积概率是5%和95%的NDVI值。本研究采用陈楠等[27]的标准进行划分,将其分为极低植被覆盖度(0~10%)、低植被覆盖度(10%~30%)、中植被覆盖度(30%~60%)、高植被覆盖度(60%~100%)。
1.3.2 转移矩阵
转移矩阵是植被覆盖度结构转变和变化方向的基础,通过转移矩阵模型能够全面刻画出各等级植被覆盖度间的时空演变过程及结构特征[28]。本研究使用Arcgis 10.2软件对2000-2020年研究区不同等级植被覆盖度进行叠加运算,后通过Excel表格来建立2000-2020年开都-孔雀河流域植被覆盖度转移矩阵。
1.3.3 Sen + Mann-Kendall趋势分析
Sen趋势度与Mann-Kendall检验结合,可以增强长时间序列趋势分析的抗噪能力,提高检验结果准确度[29]。Sen趋势度的公式为:
$ \beta =M{edian}\left (\dfrac{{{x}}_{{j}}-{{x}}_{{i}}}{{j-i}}\right){,}\forall {j} > i 。 $

(2) 式中:趋势度β判断为FVC变化趋势的升降;xi、xj为FVC中的序列数据;i和j为不同时间序列长度。若β < 0时,时间序列呈下降趋势;反之呈上升趋势。
Mann-Kendall方法是FVC时间序列趋势性检验的方法,其统计检验方法如下:
$ S = \displaystyle\sum\limits_{i = 1}^{{{n - }}1} {\displaystyle\sum\limits_{j = i + 1}^n {{sgn} \left ( {{x_j} - {x_i}} \right)} } {;} $

(3) $ {sgn}\left ({{x}}_{{j}}{{-x}}_{{i}}\right)=\left\{ {\begin{array}{*{20}{l}} { + 1{,} }&{ {x}_{j}-{x}_{i} > 0 }\\ { 0{,} }&{ {x}_{j}-{x}_{i}=0 }\\ { -1{,} }&{ {{x}}_{{j}}-{{x}}_{{i}} < 0 }\end{array}} \right.; $

(4) $ V{{ar}}\left ( S \right) = \dfrac{{{{n}}\left ( {{{n - 1}}} \right)\left ( {2{{n}} + {{5}}} \right)}}{{18}} {;} $

(5) $ Z=\left\{ {\begin{array}{*{20}{l}} {\dfrac{S-1}{\sqrt{V{ar}\left (S\right)}}{,} }&{ \left (S > 0\right) }\\ { 0{,} }&{\left (S=0\right) }\\ { \dfrac{S + 1}{V{ar}\left (S\right)}{,} }&{ \left (S < 0\right) }\end{array}} \right.。 $

(6) 式中:xi和xy为FVC序列数据,sgn为符号函数,Var为计算方差函数,θ为xj与xi的差值。依据n值大小的不同,显著性检验统计量的选取也不同。在显著性水平α下,Z1-α/2在正态分布表中所对应的值,结合β值和Z值,将植被覆盖度趋势变化的显著性分为7个等级:当β > 0,且通过99%的显著性检验时,表明植被覆盖度呈极显著上升趋势;当β > 0,且通过95%的显著性检验时,表明植被覆盖度呈显著上升趋势;当β > 0,通过90%的显著性检验时,表明植被覆盖度呈不显著上升趋势;当β = 0时,表明植被覆盖度未发生变化;同理可得植被覆盖度呈下降趋势。
1.3.4 指标选取及信息提取
1)指标选取:本研究选取高程、坡向、气温、降水、人口密度、地均GDP、牲畜年底头数、农业人口密度、土地利用类型9个驱动因子对开都-孔雀河植被覆盖度变化的影响(表1)。
表 1 驱动因子指标Table 1. Driving factors for fractional vegetation cover in the Kaidu-Kongqi River Basin类型
Category影响因子
Factor单位
Factor unit地形
Topography高程(X1)
Elevationm 坡向(X2)
Aspect气候 Climate 气温(X3)
Temperature℃ 降水(X4)
Precipitationmm 社会经济
Socioeconomic人口密度(X5)
Population densityperson·km−2 地均GDP (X6)
Average land GDPCNY·km−2 牲畜年底头数(X7)
Head count of livestock at the
end of the year104 head 农业人口密度(X8)
Agricultural population densityperson·km−2 土地利用类型(X9)
Land use type2)信息提取:地形因子、气象因子、社会经济因子按照自然间断点分级法进行重分类,分别为9类、9类、6类;土地利用类型按照中国土地利用/土地覆盖数据的分类系统,根据土地资源及其利用属性进行重分类,分为耕地、林地、草地、水域、建设用地和未利用地;而后在Arcgis 10.2软件中利用渔网工具将开孔河流域进行规则网格划分,设置网格大小为3 km × 3 km,取每个网格的中心点作为采样点,共计10 688个。再获取采样点对应各个驱动因子的数据。
1.3.5 地理探测器
地理探测器是一组统计方法,用来检测空间分布上的不均匀性,并揭示FVC变化的推动力[30]。
1)因子探测器
利用因子探测功能识别影响研究区FVC格局变化的因子,并分析其解释力。利用方程计算各因子对FVC的解释力,公式如下:
$ {{q}} = 1 - \dfrac{{\displaystyle\sum_{{{h}} = {{1}}}^{{L}} {{{{n}}_{{h}}}\sigma _{{h}}^2} }}{{{{n}}{\sigma ^{{2}}}}} = 1 - \dfrac{{SSW}}{{SST}} {;} $

(7) $ SSW={\displaystyle \displaystyle\sum _{{h}=1}^{L}{{n}}_{{h}}{\sigma }_{{h}}^{2}}{,}SST={n}{\sigma }^{2} 。 $

(8) 式中:h = 1, 2, …, L表示变量Y或因子X的数目;nh表示h层中的单位量;n表示总区域内的单位数量。SSW表示层内方差之和(within sum of square);SST表示全区总方差(total sum of squares)。q值在[0, 1],q值越大,说明自变量X对因变量Y的解释力越强。
2)交互探测器
用于识别不同因子(自变量)之间的交互作用,评估方法是首先分别计算两种影响因子X1和X2对Y的q值,比较单因子q值、双因子q值和两因子相互作用的总和,并评估双因子对因变量FVC的解释力是否有较大的有影响。
3)生态探测器
用于判断交互因子对植被覆盖度空间变化的影响是否存在显著性差异,通过F统计量检验:
$ F = \dfrac{{{N_{x{{1}}}}\left ( {{N_{x{{2}}}} - 1} \right) SS{W_{x{{1}}}}}}{{{N_{x{{2}}}}\left ( {{N_{x{{1}}}} - 1} \right) SS{W_{x{{2}}}}}} 。 $

(9) 式中:Nx1和Nx2表示这两因子的样本量;SSWx1和SSWx2分别表示由X1和X2形成的层内方差之和。
4)风险探测器
用于判定每个影响因子推进植被生长的合适范围或类型,用统计t测量:
$ {{{t}}_{{{{{\bar Y}}}_{{{h}} = {{1}}}} - {{{{\bar Y}}}_{{{h}} = {{2}}}}}} = \dfrac{{{{\bar Y}_{{{h}} = 1}} - {{\bar Y}_{{{h}} = 2}}}}{{{{\left[ {\dfrac{{V{{ar}}\left ( {{{\bar Y}_{{{h}} = 1}}} \right)}}{{{{{n}}_{{{h}} = 1}}}} + \dfrac{{Var\left ( {{{\bar Y}_{h = 2}}} \right)}}{{{n_{h = 2}}}}} \right]}^{1/2}}}} 。 $

(10) This page contains the following errors:
error on line 1 at column 1: Start tag expected, '<' not foundBelow is a rendering of the page up to the first error.
2. 结果与分析
2.1 植被覆盖度时空分布特征
基于开都-孔雀河流域NDVI数据集(2000-2020年),通过像元二分模型得出植被覆盖度。该流域植被覆盖度年际变化表明,21年来流域植被覆盖度多年均值为27.1% (图2),其中最小值出现在2001年,FVC均值为25.5%,而在2019年植被覆盖度均值达到最大值,为29.0%。21年间流域内FVC总体上呈现缓慢增长趋势,年增长速率为每10年0.013,表明该流域植被生长状况呈现轻微变好趋势。2000-2020年植被覆盖变化前期平稳后期增长:2000-2011年,植被覆盖度均值略有小幅度波动。在2011年之后,相较于前11年,植被FVC均值大幅度升高,到2020年共计增加了2.3%,说明植被活动朝增强的方向发展,有相对的提升。
![]() 图 2 开都-孔雀河流域植被覆盖度年际变化Figure 2. Interannual variation of vegetation cover in the Kaidu-Kongqi River Basin from 2000 to 2020
图 2 开都-孔雀河流域植被覆盖度年际变化Figure 2. Interannual variation of vegetation cover in the Kaidu-Kongqi River Basin from 2000 to 20202000-2020年开孔河流域植被覆盖变化势头比较明显(图3),开孔河流域植被覆盖等级呈现西北高-东南低的走向,中、高植被覆盖度分布在研究区西北的山区,低植被覆盖度主要分布在山区边缘以及中部平原,极低植被覆盖度主要分布在东南部的戈壁、荒漠区,研究区各等级植被覆盖度面积整体占比依次为极低植被覆盖度 > 高植被覆盖度 > 低植被覆盖度 > 中植被覆盖度。根据该流域2000-2020年植被覆盖面积变化(表2)可知,整个研究区的情况呈现出极低覆盖度面积减少,高、中、低植被覆盖度面积增加的趋势。极低植被覆盖度是研究区覆盖类型分布度最高的一类,约占研究区面积的50%。由于极低植被覆盖度面积主要集中在荒漠、裸地区,受国家实行生态政策的保护,极低覆盖面积变化幅度最大,减少量为5 010.31 km2,减幅为7.72%;低植被覆盖度面积呈先增后减的倒“U”型动态变化,在2010年时低覆盖度的面积较2000年的增加2.16%,其增加的面积主要出现在博斯腾湖南部,但在20年间共增加1 253.19 km2;2000-2020年高植被覆盖面积增长最多,增加量为1 993.13 km2,在2000年基础上增长了8.18%,其增加的高覆盖度面积主要分布在焉耆盆地;中覆盖度面积增长次之,较2000年增加了1 764.00 km2,增幅为16.31%,其面积增长主要位于开都河北部周边以及山谷。
![]() 图 3 2000-2020年开都-孔雀河流域植被覆盖度空间格局Figure 3. Spatial patterns of vegetation cover in the Kaidu-Kongkqi River Basin from 2000 to 2020表 2 2000-2020年开都-孔雀河流域植被覆盖度面积统计Table 2. Vegetation coverage area of the Kaidu-Kongqi River Basin from 2000 to 2020
图 3 2000-2020年开都-孔雀河流域植被覆盖度空间格局Figure 3. Spatial patterns of vegetation cover in the Kaidu-Kongkqi River Basin from 2000 to 2020表 2 2000-2020年开都-孔雀河流域植被覆盖度面积统计Table 2. Vegetation coverage area of the Kaidu-Kongqi River Basin from 2000 to 2020覆盖度类型
Coverage type2000 2010 2020 2000-2020 面积
Area/
km2比例
Percentage/
%面积
Area/
km2比例
Percentage/
%面积
Area/
km2比例
Percentage/
%变化量
Variations/
km2变化率
Rate of
change/%极低植被覆盖区
Extremely low
vegetation coverage64 941.19 56.27 60 678.69 52.58 59 930.88 51.93 −5 010.31 −7.72 低植被覆盖区
Low vegetation coverage15 282.75 13.24 17 768.94 15.40 16 535.94 14.33 1 253.19 8.20 中植被覆盖区
Medium vegetation coverage10 814.56 9.37 12 152.88 10.53 12 578.56 10.90 1 764.00 16.31 高植被覆盖区
High vegetation coverage24 365.38 21.11 24 803.38 21.49 26 358.50 22.84 1 993.13 8.18 2.2 植被覆盖时空变化特征
2.2.1 植被覆盖等级转移矩阵
2000-2020年,植被覆盖度等级之间转化频繁,以极低植被覆盖度与低植被覆盖度之间转化为主。如表3所列,2000-2020年植被覆盖等级总转移面积为16 350.94 km2,其中极低覆盖度转出面积最大为6 123.31 km2,主要转化为低植被覆盖度,转出面积为4 747.75 km2;而其他植被覆盖度等级转为极低植被覆盖的面积有1 113.00 km2,主要来自于低覆盖度1 099.00 km2。低植被覆盖度转出面积为4 442.88 km2,转出部分主要为中覆盖度、高覆盖度,面积分别占比为11.76%和10.12%;低覆盖度转入面积是2020年植被覆盖度等级中最多的,面积为5 696.06 km2。中植被覆盖度转入总量为5 043.50 km2,主要转入类型为高覆盖度,面积为2 476.81 km2。2020年高植被覆盖度总面积为26 358.50 km2,其中未发生变化的面积为21 860.13 km2,主要转入类型为中覆盖度,转入量为2 345.63 km2。
表 3 2000-2020年开都-孔雀河流域植被覆盖度转移矩阵Table 3. Vegetation coverage transition matrix in the Kaidu-Kongqi River Basin from 2000 to 2020km2 覆盖度类型
Coverage type极低植被覆盖度
Extremely low
vegetation coverage低植被覆盖度
Low vegetation
coverage中植被覆盖度
Medium vegetation
coverage高植被覆盖度
High vegetation
coverage2000年总计
Total of 2000转出
Transfer
out极低植被覆盖度
Extremely low vegetation coverage58 817.88 4 747.75 768.94 606.63 64 941.19 6 123.31 低植被覆盖度
low vegetation coverage1 099.00 10 839.88 1 797.75 1 546.13 15 282.75 4 442.88 中植被覆盖度
Medium vegetation coverage9.00 924.88 7 535.06 2 345.63 10 814.56 3 279.50 高植被覆盖度
High vegetation coverage5.00 23.44 2 476.81 21 860.13 24 365.38 2 505.25 2020年总计 Total of 2020 59 930.88 16 535.94 12 578.56 26 358.50 115 403.88 16 350.94 转入 Transfer in 1 113.00 5 696.06 5 043.50 4 498.38 2.2.2 植被覆盖度变化趋势
2000-2020年开孔河流域植被覆盖变化趋势表明,植被覆盖趋势呈无变化的面积为12 736.06 km2,约有41.31%的区域面积呈现减少趋势,还有47.66%的区域面积呈现增加趋势,说明21年来开孔河流域的植被有轻微改善趋势(图4、表4)。极显著增加的面积为12 185.69 km2,约为10.56%,主要分布在开都河中游下部以及博斯腾湖的西北部、东南部;显著与不显著增加的区域基本相同,面积分别为8 375.56和34 438.88 km2,两者分别占7.26%和29.84%,主要分布在研究区西北部和中部博斯腾湖周围;图中不显著减少区域、极显著减少和显著减少的3个区域分布均在一起,其中不显著减少的区域面积为35 799.19 km2,占研究区总面积比例最大为31.02%,主要在开都河上游的周围区域和孔雀河下游的荒漠区分布;极显著减少的区域面积为4 939.31 km2,其主要在流域东南部分布较多,在研究区北部分布较少,而显著减少区域伴随着极显著减少区域分布,极显著区域面积占比最小为4.28%。
![]() 图 4 2000-2020年开都-孔雀河流域植被覆盖度变化趋势Figure 4. Vegetation coverage trends in the Kaidu-Kongqi River Basin from 2000 to 2020表 4 2000-2020年开都-孔雀河流域植被覆盖度变化趋势统计Table 4. Statistical analysis of vegetation coverage trend changes in the Kaidu-Kongqi River Basin from 2000 to 2020
图 4 2000-2020年开都-孔雀河流域植被覆盖度变化趋势Figure 4. Vegetation coverage trends in the Kaidu-Kongqi River Basin from 2000 to 2020表 4 2000-2020年开都-孔雀河流域植被覆盖度变化趋势统计Table 4. Statistical analysis of vegetation coverage trend changes in the Kaidu-Kongqi River Basin from 2000 to 2020变化趋势等级
Change trend level置信度
Confidence/%面积
Area/km2比例
Percentage/%极显著增加
Extremely significant
increase99 12 185.69 10.56 显著增加
Significant increase95 8 375.56 7.26 不显著增加
Non-significant increase90 34 438.88 29.84 无变化 No change − 12 736.06 11.04 不显著减少
Non-significant decrease90 35 799.19 31.02 显著减少
Significant decrease95 6 929.19 6.00 极显著减少
Extremely significant
decrease99 4 939.31 4.28 2.3 驱动因素的空间异质性分析
2.3.1 探测因子影响力分析
各驱动因子对植被覆盖变化的影响强弱可以通过因子探测得出的q值大小来阐明。因子影响力结果表明,各驱动因子的q值排序为高程(0.481) > 土地利用类型(0.472) > 气温(0.430) > 农业人口密度(0.389) = 人口密度(0.389) > 降水(0.385) > 牲畜年底头数(0.381) > 地均GDP (0.277) > 坡向(0.005)(图5)。 因此,影响流域植被覆盖度空间分层差异的主要影响因子是高程和土地利用类型,两个因子的解释力均超过0.45;气温、农业人口密度、人口密度、降水及牲畜年底头数作为次级影响因子,解释力均在0.35以上;地均GDP的解释力超过0.20;坡向的单个因素解释力最小。
![]() 图 5 各驱动因子解释力X1:高程;X2:坡向;X3:气温;X4:降水;X5:人口密度;X6:地均GDP;X7:牲畜年底头数;X8:农业人口密度;X9:土地利用类型。下同。Figure 5. Explanatory power of each driving factorX1: elevation; X2: aspect; X3: temperature; X4: precipitation; X5: population density; X6: average land GDP; X7: head count of livestock at the end of the year; X8: agricultural population density; X9: land use type. This is applicable for the following tables and figures as well.
图 5 各驱动因子解释力X1:高程;X2:坡向;X3:气温;X4:降水;X5:人口密度;X6:地均GDP;X7:牲畜年底头数;X8:农业人口密度;X9:土地利用类型。下同。Figure 5. Explanatory power of each driving factorX1: elevation; X2: aspect; X3: temperature; X4: precipitation; X5: population density; X6: average land GDP; X7: head count of livestock at the end of the year; X8: agricultural population density; X9: land use type. This is applicable for the following tables and figures as well.2.3.2 探测交互作用分析
交互探测器主要是评估两因子共同作用时是否会增加或减少对植被覆盖度的解释力,所有因子两两交互都增强了对FVC变化的贡献力,其中除了坡向与其他因子的交互作用呈非线性增强,其余均呈现双因子增强(图6)。土地利用类型与除坡向和地均GDP的其他驱动因子的交互作用,解释力在0.65以上,分别为X1∩X9 (0.743)、X3∩X9 (0.727)、X4∩X9 (0.706)、X5∩X9 (0.663)、X7∩X9 (0.662)、X8∩X9 (0.658),从解释力角度看,土地利用类型与其他驱动因子的协同作用下,显著增强了土地利用类型对植被覆盖度的影响;解释力在0.50以上的有X1∩X3 (0.584)、X1∩X4 (0.554)、X1∩X5 (0.599)、X1∩X6 (0.592)、X1∩X7 (0.606)、X1∩X8 (0.603)、X3∩X5 (0.512)、X3∩X6 (0.510)、X3∩X7 (0.510)、X3∩X8 (0.502)、X4∩X5 (0.503),由此看出,高程、气温分别与各因子之间交互作用对植被覆盖度的影响较为强烈,同时坡向与各驱动因子之间交互作用比坡向对植被覆盖度的直接影响较大,说明两因子的叠加效果远大于单一因子对植被覆盖的影响。
2.3.3 探测显著性差异分析
生态因子检测用于判断交互因子在植被FVC的空间分布上有无显著性差异,由探测因子显著性分析结果(表5)可知,高程和其他因子之间差异不显著(P > 0.05);坡向与除高程的其余驱动因素对FVC空间分布均具有显著差异(P < 0.05);气温和坡向、土地利用类型之间存在显著性差异,与其他因子无显著性差异;降水与高程、气温、人口密度、地均GDP、牲畜年底头数、农业人口密度之间的共同作用对植被覆盖变化无显著性差异,与土地利用类型和坡向对植被覆盖度空间变化有显著差异;人口密度与坡向、土地利用类型的相互作用对植被覆盖度空间分布存在有显著性差异;地均GDP分别与高程、气温、降水、人口密度的交互作用对植被覆盖时空变化存在无显著性差异;牲畜年底头数和农业人口密度分别与坡向、地均GDP、土地利用类型有显著差异,而与其他因素无显著差异;土地利用类型与高程的叠加作用下对植被覆盖变化不存在显著性差异,而与其他因子均具有显著差异。
表 5 探测因子显著性分析(置信水平95%)Table 5. Significance analysis of each detected factor (Confidence level 95%)因子 Factor X1 X2 X3 X4 X5 X6 X7 X8 X9 X1 X2 N X3 N Y X4 N Y N X5 N Y N N X6 N Y N N N X7 N Y N N N Y X8 N Y N N N Y N X9 N Y Y Y Y Y Y Y N表示在0.05水平上交互因子叠加对FVC变化的影响不存在显著性差异,Y表示有显著性差异。
N: no significant difference in the influence of interaction factors on FVC change at the 0.05 level; Y: significant difference at the 0.05 level.2.3.4 探测因子适宜性分析
不同驱动因子的植被FVC均值差异明显(表6)。随高程、降水、人口密度的增加,研究区内植被覆盖度增高,分别在2 932~3 291 m、228.11~270.46 mm、1 人·km−2分区时,FVC达到最大,分别为79.8%、85.0%和61.9%;而随着气温升高,植被覆盖度呈下降趋势,在–0.73~2.26 ℃的适宜范围内FVC达到最大(85.3%),说明这个温度范围促进了植被生长;FVC随地均GDP、牲畜年底头数、农业人口密度的增加呈先增高后降低的趋势和波动变化,分别在99 735~257 254 元·km−2、43.1万~116万头、1~3人·km−2分区时,FVC达到最大,分别为50.7%、61.9%和61.9%;坡向为北坡时最适宜植被生长,植被覆盖度达到36.6%;土地利用类型为耕地时,FVC达到最大(78.9%),这可能由于人为开垦荒地,导致耕地面积迅速扩张,使得植被覆盖面积也随之增加。
表 6 驱动因子的适宜类型或范围(置信水平95%)Table 6. Appropriate type or range of driver (Confidence level 95%)驱动因子
Driving factor适宜类型或范围
Suitable type or range植被覆盖度
Fractional vegetation cover/%高程 Elevation/m 2 932~3 291 79.8 坡向 Aspect 北坡 North slope 36.6 气温 Temperature/℃ −0.73~2.26 85.3 降水 Precipitation/mm 228.11~270.46 85.0 人口密度 Population density/(person·km−2) 1 61.9 地均GDP Average land GDP/(CNY·km−2) 99 735~257 254 50.7 牲畜年底头数 Head count of livestock at the end of the year/(×104 head) 43.1~116.0 61.9 农业人口密度 Agricultural population density/(person·km−2) 1~3 61.9 土地利用类型 Land use type 耕地 Cultivated land 78.9 3. 讨论
开都-孔雀河流域植被覆盖等级呈西北高-东南低趋势,2000-2020年流域植被生长状况整体呈增长态势,这与万洪秀等[31]研究结果一致。21年间开孔河流域FVC空间上发生明显变化,巴州曾存在草地退化、过度放牧等生态问题,2011年巴州贯彻国家草原生态保护补助奖励机制和禁牧、轮牧、休牧、草蓄平衡制度[32],植被覆盖度持续性增加。其中库尔勒市周围区域植被增加尤为明显,主要与耕地扩增有关[33];近年来降水量增加、蒸发量减少[34]使焉耆盆地和博斯腾湖南部区域的植被覆盖度显著增加;山区降水量减少,气温降低和平原区气温升高,降水减少[35],是导致开都河东部和孔雀河南部植被退化的主要原因。由此得出,研究区植被覆盖变化整体呈现轻微改善的趋势,但仍然存在较大的生态环境问题。
本研究中,高程和土地利用类型是影响开都-孔雀河流域植被覆盖度变化的主要驱动因子,气温对植被覆盖度的影响次之,解释力均在0.40以上,并且影响力表现为高程 > 土地利用类型 > 气温。高程对植被覆盖变化有着不可忽视的作用,王涛等[36]研究高程对博斯腾湖流域植被变化影响,发现随着海拔高度增加,植被生长影像呈正态分布。研究区地势落差大,西北部多为山地,海拔较高,主要表现为高、中植被覆盖度;中部和东南部多以平原、荒漠为主,海拔较低且蒸发量大,主要表现为低、极低植被覆盖度。人类活动主要通过改变土地利用类型来影响植被变化,研究期间流域内耕地面积增加较多,其主要分布在库尔勒市区周边以及焉耆盆地[37],植被覆盖度也随之增加,其中高程与土地利用的交互作用对植被覆盖度时空变化的解释力最大。植被对气候变化较为敏感[38],尤其在新疆干旱、半干旱区域,降水稀少且蒸发剧烈,但在气温逐渐升高情况下,相较于降水,气温成为限制植被长势的重要因子[39]。因此,合理地研析和阐明驱动因素及其因子之间的协同作用对植被时空变化的影响,探索驱动因素促进植被生长的适宜范围,从而有助于当地政府在有利范围内干预植被变化,减轻人类发展带来的负面影响,有效地指导植被恢复和保护。
本研究仅对年际FVC均值变化进行讨论,尚未对年内FVC均值变化,以及未量化政策对植被变化的影响进行研究。在今后的工作中,将深刻探讨开都-孔雀河流域对自然及人类驱动因素的响应,详细阐明植被覆盖度时空变化特征,并对植被变化的驱动机制进行系统地解释。
4. 结论
1) 2000-2020年开都-孔雀河流域植被覆盖度呈缓慢增加趋势,上升幅度为0.013·10 a−1,2000-2011年植被覆盖度变化较为平稳,2011年之后,植被覆盖度持续增加。空间上,植被覆盖度呈现从西北向东南递减的空间分布特征。在20年间高植被覆盖度面积增加1 993.13 km2,主要分布在库尔勒市周边区域和焉耆盆地。
2)开都-孔雀河流域以极低覆盖度与低覆盖度之间转化为主,其中极低植被覆盖度转出面积远大于转入面积,主要转化为低覆盖度。开孔河流域植被覆有轻微的改善趋势,其中17.82%的区域呈极显著、显著增加趋势;10.28%的区域呈极显著、显著减少趋势;71.90%的区域呈不显著和无变化趋势。
3)高程和土地利用类型是造成区域内植被分布格局的主要因素,坡向与流域植被覆盖变化的相关性最弱,其中高程和土地利用类型两两相交的解释力最高为0.743,除了土地利用与坡向的相互作用,与其他驱动因子的共同作用下解释力均超过0.50,其次高程、气温与其余因子之间交互作用对植被覆盖度的影响较为强烈。
4)风险因子探测分析发现促进植被生长的各主要因子最适宜范围:气温为–0.73~2.26 ℃,降水为228.11~270.46 mm,高程为2 932~3 291 m,坡向为北坡适合开孔河流域植被生长,人口密度为1人·km−2,地均GDP为99 735~257 254 元·km−2,牲畜年底头数为43.1万~116万头,农业人口密度为1~3人·km−2,土地利用类型为耕地,植被覆盖度最大。该成果为该流域植被生长变化的驱动因素分析提供了参考和借鉴。
参考文献
[1] STERNER R W, ELSER J J. Ecological Stoichiometry: The Biology of Elements From Molecules to The Biosphere. Princeton: Princeton University Press, 2002: 225-226.
[2] GIORDANO M. Homeostasis: An underestimated focal point of ecology and evolution. Plant Science, 2013, 211: 92-101. doi: 10.1016/j.plantsci.2013.07.008
[3] 张婷婷, 刘文耀, 黄俊彪, 胡涛, 汤丹丹, 陈泉. 植物生态化学计量内稳性特征. 广西植物, 2019, 39(5): 701-712. ZHANG T T, LIU W Y, HUANG J B, HU T, TANG D D, CHEN Q. Characteristics of plant ecological stoichiometry homeostasis. Guihaia, 2019, 39(5): 701-712.
[4] GUSEWELL S. N ꞉ P ratios in terrestrial plants: Variation and functional significance. New Phytologist, 2004, 164: 243-266.
[5] 蒋利玲, 曾从盛, 邵钧炯, 周旭辉. 闽江河口入侵种互花米草和本地种短叶茳芏的养分动态及植物化学计量内稳性特征. 植物生态学报, 2017, 41(4): 450-460. doi: 10.17521/cjpe.2016.0193 JIANG L L, ZENG C S, SHAO J J, ZHOU X H. Plant nutrient dynamics and stoichiometric homeostasis of invasive species Spartina alterniflora and native Cyperus malaccensis var. brevifolius in the Minjiang River estuarine wetlands. Chinese Journal of Plant Ecology, 2017, 41(4): 450-460. doi: 10.17521/cjpe.2016.0193
[6] YU Q, CHEN Q S, ELSER J J, HE N P, WU H H, ZHANG G M, WU J G, BAI Y F, HAN X G. Linking stoichiometric homeostasis with ecosystem structure, functioning, and stability. Ecology Letters, 2010, 13: 1390. doi: 10.1111/j.1461-0248.2010.01532.x
[7] YU Q, WILCOX K, PIERRE K L, KNAPP A K, HAN X, SMITH M D. Stoichiometric homeostasis predicts plant species dominance, temporal stability, and responses to global change. Ecology, 2015, 96(9): 2328-2335. doi: 10.1890/14-1897.1
[8] SU H, WU Y, XIA W, YANG L, CHEN J, HAN W, FANG J, XIE P. Stoichiometric mechanisms of regime shifts in freshwater ecosystem. Water Research, 2019, 149: 302-310. doi: 10.1016/j.watres.2018.11.024
[9] RAO QY, SU H J, RUAN L W, DENG X W, WANG L T, RAO X, LIU J R, XIA W L, XU P K, SHEN H, CHEN J, XIE P. Stoichiometric and physiological mechanisms that link hub traits of submerged macrophytes with ecosystem structure and functioning. Water Research, 2021, 202: 117-392.
[10] 王萨仁娜. 3S技术支持下的荒漠草原不同载畜率水平植被变化研究. 呼和浩特: 内蒙古农业大学硕士学位论文, 2016. WANG S R N. Study on vegetation change and mechanism under different stocking rate in desert steppe based on 3S technology. Master Thesis. Hohhot: Inner Mongolia Agricultural University, 2016.
[11] HESSEN D O, AGREN G I, ANDERSON T R, ELSER J J, RUTIER P C. Carbon sequestration in ecosystems: The role of stoichiometry. Ecology, 2004, 85: 1179-1192. doi: 10.1890/02-0251
[12] YU Q, ELSER J J, HE N P, WU H H, CHEN Q S, ZHANG G M, HAN X G. Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland. Oecologia, 2011, 166(1): 1-10. doi: 10.1007/s00442-010-1902-z
[13] GU Q, ZAMIN T J, GROGAN P. Stoichiometric homeostasis: A test to predict tundra vascular plant species and community-level responses to climate change. Arctic Science, 2017, 3(2): 320-333. doi: 10.1139/as-2016-0032
[14] 鲍士旦. 土壤农化分析(第三版). 北京: 中国农业出版社, 2000. BAO S D. Soil and Agricultural Chemistry Analysis (Third Edition). Beijing: China Agriculture Press, 2000.
[15] PERSSON J, FINK P, GOTO A. To be or not to be what you eat: Regulation of stoichiometric homeostasis among autotrophs and heterotrophs. Oikos, 2010, 119: 741-751. doi: 10.1111/j.1600-0706.2009.18545.x
[16] VITOUSEK P M, PORDER S, HOULTON B Z. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen–phosphorus interactions. Ecological Applications, 2010, 20(1): 5-15. doi: 10.1890/08-0127.1
[17] SISTLA S A, APPLING A P, LEWANDOWSKA A M, TAYLOR B N, WOLF A A. Stoichiometric flexibility in response to fertilization along gradients of environmental and organismal nutrient richness. Oikos, 2015(124): 949-959.
[18] PENG H Y, CHEN Y H, YAN Z B, HAN W X. Stage-dependent stoichiometric homeostasis and responses of nutrient resorption in Amaranthus mangostanus to nitrogen and phosphorus addition. Scientific Reports, 2016, 6: 37219. doi: 10.1038/srep37219
[19] 金晓明, 于良斌, 张颖琪, 王秋红. 群落演替对呼伦贝尔草地两种优势植物繁殖分配及生态化学计量的影响. 应用生态学报, 2020, 31(3): 787-793. JING X M, YU L B, ZHANG Y Q, WANG Q H. Effects of community succession on plant reproductive allocation and ecological stoichiometry for two dominant species in the Hulunbuir Grassland, China. Chinese Journal of Applied Ecology, 2020, 31(3): 787-793.
[20] 赵君, 张仁彭, 李新娥, 陈凌云, 王刚. 甘南亚高寒草甸金露梅叶片氮磷化学计量学动态. 兰州大学学报, 2011, 47(2): 88-92. ZHAO J, ZHANG R P, LI X E, CHEN L Y, WANG G. Leaf nitrogen and phosphorus stoichiometry in Potentilla fruticosa L. affected by internal and external factors in the subalpine in Gannan. Journal of Lanzhou University, 2011, 47(2): 88-92.
[21] 宁志英, 李玉霖, 杨红玲, 张子谦. 科尔沁沙地优势固沙灌木叶片氮磷化学计量内稳性. 植物生态学报, 2019, 43(1): 46-54. doi: 10.17521/cjpe.2018.0100 NING Z Y, LI Y L, YANG H L, ZHANG Z Q. Nitrogen and phosphorus stoichiometric homoeostasis in leaves of dominant sand-fixing shrubs in Horqin Sandy Land, China. Chinese Journal of Plant Ecology, 2019, 43(1): 46-54. doi: 10.17521/cjpe.2018.0100
[22] FRIEDMAN J. The evolution of annual and perennial plant life histories: ecological correlates and genetic mechanisms. Annual Review of Ecology, Evolution, and Systematics, 2020, 51: 461-481. doi: 10.1146/annurev-ecolsys-110218-024638
[23] 瞿王龙, 杨小鹏, 张存涛, 魏冰. 干旱、半干旱地区天然草原灌木及其肥岛效应研究进展. 草业学报, 2015, 24(4): 201-207. QU W L, YANG X P, ZHANG C T, WEI B. Shrub-mediated “fertile island” effects in arid and semi-arid grassland. Acta Prataculture Sinica, 2015, 24(4): 201-207.
[24] LI Y, ZHOU X, ZHANG Y. Shrub modulates the stoichiometry of moss and soil in desert ecosystems, China. Journal of Arid Land, 2019, 11(4): 579-594. doi: 10.1007/s40333-019-0057-y
[25] NING Z, LI Y, ZHAO X Y, HAN D, ZHANG J. Comparison of leaf and fine root traits between annuals and perennials, implicating the mechanism of species changes in grassland. Frontiers in Plant Science, 2022, 12: 547-778.
[26] 苏强. 浮游动物化学计量学稳态性特征研究进展. 生态学报, 2012, 32(22): 7213-7219. doi: 10.5846/stxb201110141522 SU Q. The framework of stoichiometry homeostasis in zooplankton elemental composition. Acta Ecologica Sinica, 2012, 32(22): 7213-7219. doi: 10.5846/stxb201110141522
[27] 严正兵, 金南瑛, 韩廷申, 方精云. 氮磷施肥对拟南芥叶片碳氮磷化学计量特征的影响. 植物生态学报, 2013, 37(6): 551-557. doi: 10.3724/SP.J.1258.2013.00056 YAN Z B, JIN N Y, HAN T S, FANG J Y. Effects of nitrogen and phosphorus fertilization on leaf carbon, nitrogen and phosphorus stoichiometry of Arabidopsis thaliana. Chinese Journal of Plant Ecology, 2013, 37(6): 551-557. doi: 10.3724/SP.J.1258.2013.00056
[28] EREL R, ANNETTE B, CAPOWIEZ L, DOUSSAN C, ARNAL D, SOUCHE G, GAVALAND A, FRITZ C, ERIC J W, SALVI S V, MARIE L C, HUND A, HINSIGER P. Soil type determines how root and rhizosphere tratis relate to phosphorus acquisition in field-grown mazie genotypes. Plant and Soil, 2017, 412(1): 115-132.
[29] 王彦丽, 范庭, 王旭东, 张育林, 李利敏, 郑洺钧. 长期磷肥不均投入下黄土高原土壤磷素有效性及与土壤性质关系分析. 水土保持学报, 2019, 33(5): 237-242, 250. WANG Y L, FAN T, WANG X D, ZHANG Y L, LI L M, ZHENG M J. Analysis of phosphorus availability and its relationship with soil properties in the Loes Plateau under long-term lmbalanced phosphorus input. Journal of Soil and Water Conservation, 2019, 33(5): 237-242, 250.
[30] 靖淑慧, 刘加珍, 陈永金, 张天举. 滨海湿地不同群落对土壤磷的有效性影响研究. 土壤通报, 2018, 49(2): 392-401. JING S H, LIU J Z, CHEN Y J, ZHANG T J. Effects of plant communities on soil phosphorus availability in coastal wetlands. Chinese Journal of Soil Science, 2018, 49(2): 392-401.
[31] ZHANG K, SU Y, YANG R. Variation of soil organic carbon, nitrogen, and phosphorus stoichiometry and biogeographic factors across the desert ecosystem of Hexi Corridor, northwestern China. Journal of Soils and Sediments, 2019, 19(1): 49-57. doi: 10.1007/s11368-018-2007-2
[32] 孙临泉, 陈子学, 张洪立, 刘冰, 刘丽丽. 微生物植酸酶对土壤有机磷组分含量及有效性的影响. 天津师范大学学报(自然科学版), 2011, 31(2): 86-90. SUN L Q, CHEN Z X, ZHANG H L, LIU B, LIU L L. Effects of microbial phytase on soil organic phosphorus component and its availability. Journal of Tianjin Normal University (Natural Science Edition), 2011, 31(2): 86-90.
[33] RICHARDSON A E, SIMPSON R J. Soil microorganisms mediating phosphorus availability update on microbial phosphorus. Plant Physiology, 2011, 156(3): 989-996. doi: 10.1104/pp.111.175448
-
![]()
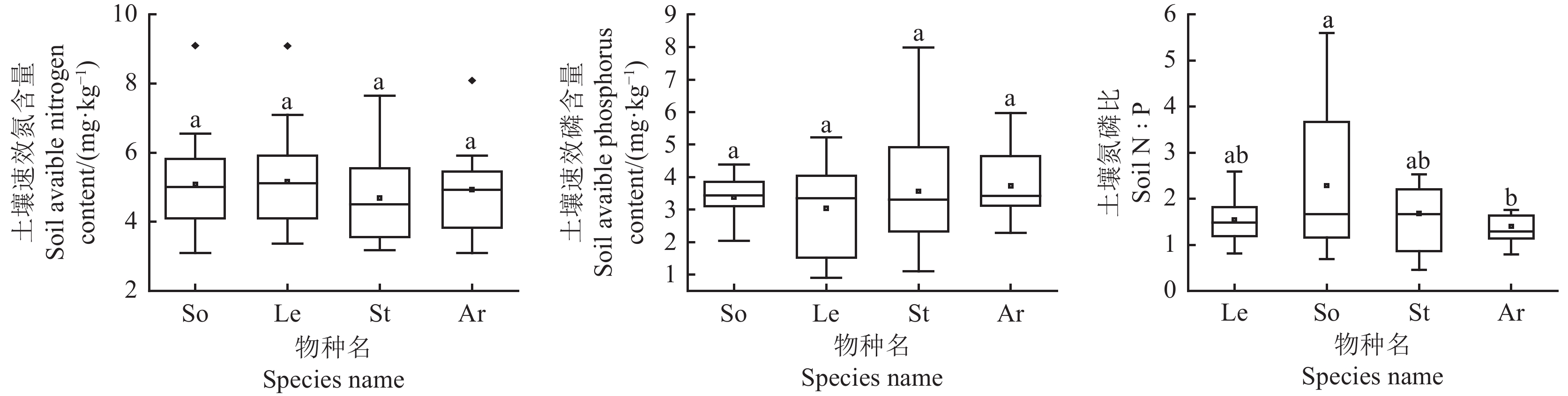
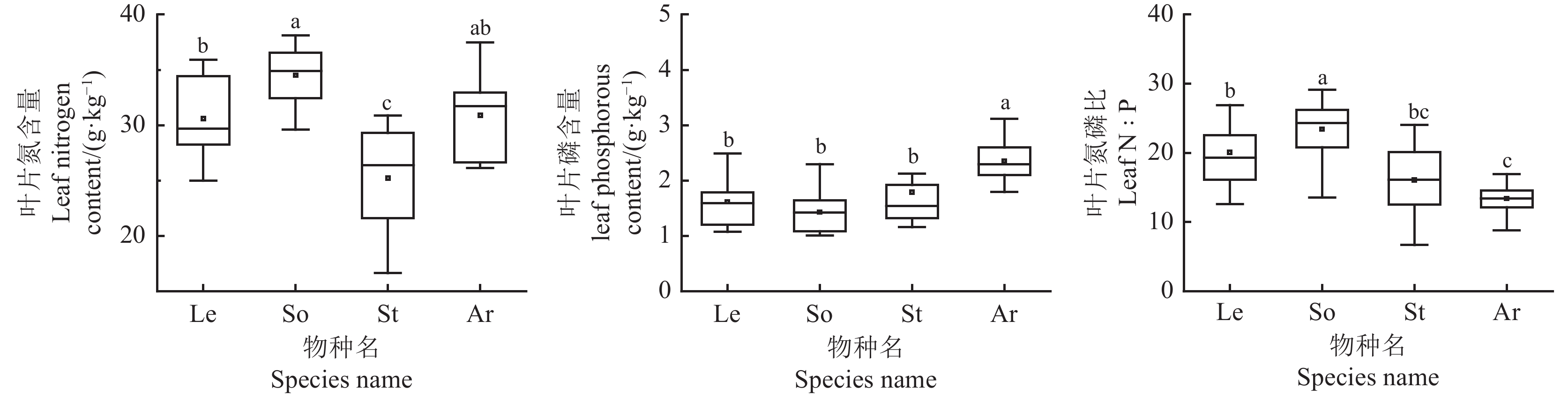
图 1 荒漠草原4种优势植物叶片N、P化学计量特征
Le,牛枝子;So ,苦豆子;St,短花针茅;Ar,猪毛蒿;不同小写字母表示差异显著(P < 0.05);下表同。
Figure 1. N and P stoichiometry characteristics of four dominant plants in a desert steppe
Le, Lespedeza potaninii; So, Sophora alopecuroides; St, Stipa breviflora; Ar, Artemisia scoparia. Different lowercase letters indicate significanct differences at the 0.05 level; This is applicable for the following table as well.
![]()
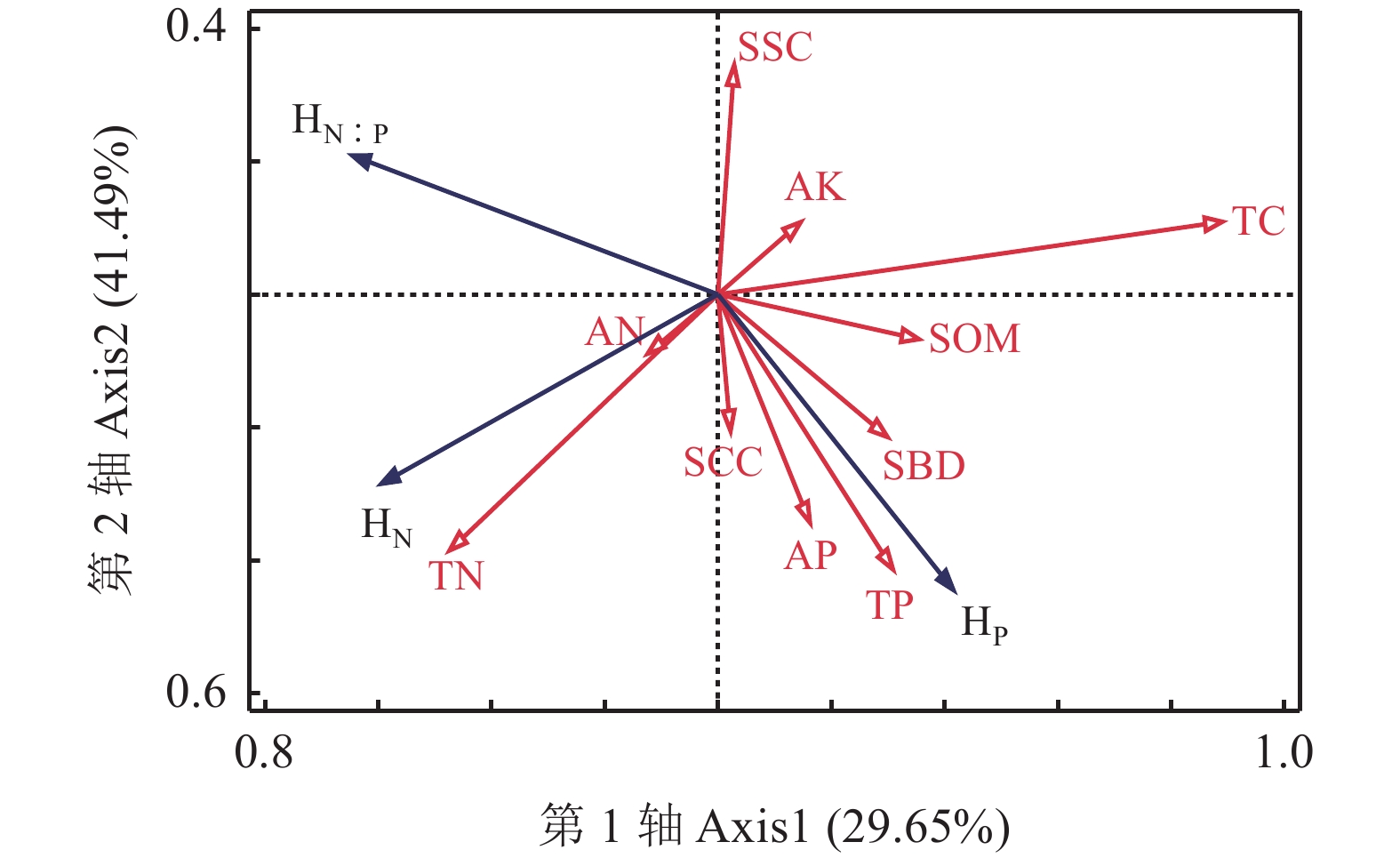
图 3 土壤理化因子对植物叶片化学计量内稳性的影响
HN,物种N内稳性;HP,物种P内稳性;HN ꞉ P,N ꞉ P内稳性;SOM, 有机质;AN,碱解氮;TN,全氮;SSC,土壤砂粒含量;SCC,黏粒含量;AP,速效磷;AK,速效钾;TP,全磷; SBD,容重。
Figure 3. Effects of soil physicochemical factors on the stoichiometric homeostasis of plant leaves
HN, N internal stability of species; HP, P internal stability of species; HN ꞉ P, N ꞉ P internal stability. SOM, soil organic matter; AN, alkaline hydrolyzable nitrogen; TN, total nitrogen; SSC, soil sandy content; SCC, soil clay content; AP, available phosphorous; AK, available potassium; TP, total phosphorous; SBD, soil bulk density.
表 1 荒漠草原4种优势植物概况
Table 1 General situation of four dominant plants in desert steppe
物种
Species科属
Family and
genus生活型
Life
form海拔
Altitude/
m盖度
Cover
degree/%密度
Density/
(plant·m−2)重要值
Importance
value样地数
Number of
plots牛枝子
Lespedeza
potaninii豆科胡枝子属
Leguminosae
Lespedeza多年生半灌木
Perennial
subshrub1 360.94 8.80 ± 0.89 10.10 ± 1.48 0.37 ± 0.13 20 苦豆子
Sophora
alopecuroides豆科槐属
Leguminosae
Styphnolobium多年生草本
Perennial
herb1 359.65 7.88 ± 1.66 2.46 ± 1.38 0.49 ± 0.31 14 短花针茅
Stipa
breviflora禾本科针茅属
Poaceae
Stipa多年生草本
Perennial
herb1 363.71 10.54 ± 0.13 8.67 ± 1.74 0.38 ± 0.26 20 猪毛蒿
Artemisia
scoparia菊科蒿属
Asteraceae
Artemisia一年生草本
Annual
herb1 345.05 22.05 ± 2.22 154.94 ± 1.69 0.27 ± 0.28 8  下载: 导出CSV
下载: 导出CSV
表 3 土壤环境变量解释的重要性排序与显著性检验结果
Table 3 Importance ranking and significance test results of soil environmental variables interpretation
土壤理化因子
Soil physicochemical factors解释量
Explains/%贡献量
Contribution/%F P 重要性排序
Important ranking全碳 Organic carbon 23.7 56.9 18.0 0.002 1 全氮 Total nitrogen 4.1 9.9 3.2 0.012 2 容重 Bulk density 2.6 6.3 2.1 0.206 3 速效磷 Available phosphorous 3.1 7.4 2.5 0.084 4 全磷 Total phosphorous 1.7 3.9 1.4 0.248 5 砂粒含量 Silt content 1.7 4.0 1.4 0.244 6 碱解氮 Available nitrogen 1.7 4.0 1.4 0.258 7 有机质 Organic matter 1.2 2.8 1.0 0.356 8 速效钾 Available potassium 0.6 1.4 0.5 0.576 9 黏粒含量 Clay content 0.3 0.8 0.3 0.728 10
下载: 导出CSV
表 2 荒漠草原4优势植物叶片N、P内稳性指数
Table 2 Stoichiometric homoeostasis indexes for four dominant plants in a desert steppe
物种
Species指标
Index内稳性指数(H)
Homeostasis indexR2 P 拟合方程
Fitted equation类型
Type牛枝子
Lespedeza
potaniniiN 5.88 0.11 0.03 lg y = 1.36 + 0.17lg x 稳态型 Homeostasis P 3.84 0.15 0.02 lg y = 0.05 + 0.26lg x 弱稳态 Weak homeostasis N ꞉ P 3.33 0.12 0.02 lg y = 1.21 + 0.30lg x 弱稳态 Weak homeostasis 苦豆子
Sophora
alopecuroidesN 3.70 0.11 0.03 lg y = 1.33 + 0.27lg x 弱稳态 Weak homeostasis P 4.00 0.62 0.05 lg y = 0.07 + 0.25lg x 弱稳态 Weak homeostasis N ꞉ P 2.94 0.19 0.02 lg y = 1.26 + 0.27lg x 弱稳态 Weak homeostasis 短花针茅
Stipa
brevifloraN 3.13 0.20 0.04 lg y = 0.58 + 0.32lg x 弱稳态 Weak homeostasis P 2.84 0.17 0.03 lg y = 0.19 + 0.35lg x 弱稳态 Weak homeostasis N ꞉ P 3.13 0.13 0.05 lg y = 0.32 + 0.26lg x 弱稳态 Weak homeostasis 猪毛蒿
Artemisia
scopariaN 2.86 0.19 0.06 lg y = 1.22 + 0.35lg x 弱稳态 Weak homeostasis P 5.88 0.10 0.04 lg y = 0.30 + 0.17lg x 稳态型 Homeostasis N ꞉ P 1.52 0.10 0.09 lg x = 0.53 + 0.66lg x 弱敏感型 Plastic
下载: 导出CSV
-
[1] STERNER R W, ELSER J J. Ecological Stoichiometry: The Biology of Elements From Molecules to The Biosphere. Princeton: Princeton University Press, 2002: 225-226.
[2] GIORDANO M. Homeostasis: An underestimated focal point of ecology and evolution. Plant Science, 2013, 211: 92-101. doi: 10.1016/j.plantsci.2013.07.008
[3] 张婷婷, 刘文耀, 黄俊彪, 胡涛, 汤丹丹, 陈泉. 植物生态化学计量内稳性特征. 广西植物, 2019, 39(5): 701-712. ZHANG T T, LIU W Y, HUANG J B, HU T, TANG D D, CHEN Q. Characteristics of plant ecological stoichiometry homeostasis. Guihaia, 2019, 39(5): 701-712.
[4] GUSEWELL S. N ꞉ P ratios in terrestrial plants: Variation and functional significance. New Phytologist, 2004, 164: 243-266.
[5] 蒋利玲, 曾从盛, 邵钧炯, 周旭辉. 闽江河口入侵种互花米草和本地种短叶茳芏的养分动态及植物化学计量内稳性特征. 植物生态学报, 2017, 41(4): 450-460. doi: 10.17521/cjpe.2016.0193 JIANG L L, ZENG C S, SHAO J J, ZHOU X H. Plant nutrient dynamics and stoichiometric homeostasis of invasive species Spartina alterniflora and native Cyperus malaccensis var. brevifolius in the Minjiang River estuarine wetlands. Chinese Journal of Plant Ecology, 2017, 41(4): 450-460. doi: 10.17521/cjpe.2016.0193
[6] YU Q, CHEN Q S, ELSER J J, HE N P, WU H H, ZHANG G M, WU J G, BAI Y F, HAN X G. Linking stoichiometric homeostasis with ecosystem structure, functioning, and stability. Ecology Letters, 2010, 13: 1390. doi: 10.1111/j.1461-0248.2010.01532.x
[7] YU Q, WILCOX K, PIERRE K L, KNAPP A K, HAN X, SMITH M D. Stoichiometric homeostasis predicts plant species dominance, temporal stability, and responses to global change. Ecology, 2015, 96(9): 2328-2335. doi: 10.1890/14-1897.1
[8] SU H, WU Y, XIA W, YANG L, CHEN J, HAN W, FANG J, XIE P. Stoichiometric mechanisms of regime shifts in freshwater ecosystem. Water Research, 2019, 149: 302-310. doi: 10.1016/j.watres.2018.11.024
[9] RAO QY, SU H J, RUAN L W, DENG X W, WANG L T, RAO X, LIU J R, XIA W L, XU P K, SHEN H, CHEN J, XIE P. Stoichiometric and physiological mechanisms that link hub traits of submerged macrophytes with ecosystem structure and functioning. Water Research, 2021, 202: 117-392.
[10] 王萨仁娜. 3S技术支持下的荒漠草原不同载畜率水平植被变化研究. 呼和浩特: 内蒙古农业大学硕士学位论文, 2016. WANG S R N. Study on vegetation change and mechanism under different stocking rate in desert steppe based on 3S technology. Master Thesis. Hohhot: Inner Mongolia Agricultural University, 2016.
[11] HESSEN D O, AGREN G I, ANDERSON T R, ELSER J J, RUTIER P C. Carbon sequestration in ecosystems: The role of stoichiometry. Ecology, 2004, 85: 1179-1192. doi: 10.1890/02-0251
[12] YU Q, ELSER J J, HE N P, WU H H, CHEN Q S, ZHANG G M, HAN X G. Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland. Oecologia, 2011, 166(1): 1-10. doi: 10.1007/s00442-010-1902-z
[13] GU Q, ZAMIN T J, GROGAN P. Stoichiometric homeostasis: A test to predict tundra vascular plant species and community-level responses to climate change. Arctic Science, 2017, 3(2): 320-333. doi: 10.1139/as-2016-0032
[14] 鲍士旦. 土壤农化分析(第三版). 北京: 中国农业出版社, 2000. BAO S D. Soil and Agricultural Chemistry Analysis (Third Edition). Beijing: China Agriculture Press, 2000.
[15] PERSSON J, FINK P, GOTO A. To be or not to be what you eat: Regulation of stoichiometric homeostasis among autotrophs and heterotrophs. Oikos, 2010, 119: 741-751. doi: 10.1111/j.1600-0706.2009.18545.x
[16] VITOUSEK P M, PORDER S, HOULTON B Z. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen–phosphorus interactions. Ecological Applications, 2010, 20(1): 5-15. doi: 10.1890/08-0127.1
[17] SISTLA S A, APPLING A P, LEWANDOWSKA A M, TAYLOR B N, WOLF A A. Stoichiometric flexibility in response to fertilization along gradients of environmental and organismal nutrient richness. Oikos, 2015(124): 949-959.
[18] PENG H Y, CHEN Y H, YAN Z B, HAN W X. Stage-dependent stoichiometric homeostasis and responses of nutrient resorption in Amaranthus mangostanus to nitrogen and phosphorus addition. Scientific Reports, 2016, 6: 37219. doi: 10.1038/srep37219
[19] 金晓明, 于良斌, 张颖琪, 王秋红. 群落演替对呼伦贝尔草地两种优势植物繁殖分配及生态化学计量的影响. 应用生态学报, 2020, 31(3): 787-793. JING X M, YU L B, ZHANG Y Q, WANG Q H. Effects of community succession on plant reproductive allocation and ecological stoichiometry for two dominant species in the Hulunbuir Grassland, China. Chinese Journal of Applied Ecology, 2020, 31(3): 787-793.
[20] 赵君, 张仁彭, 李新娥, 陈凌云, 王刚. 甘南亚高寒草甸金露梅叶片氮磷化学计量学动态. 兰州大学学报, 2011, 47(2): 88-92. ZHAO J, ZHANG R P, LI X E, CHEN L Y, WANG G. Leaf nitrogen and phosphorus stoichiometry in Potentilla fruticosa L. affected by internal and external factors in the subalpine in Gannan. Journal of Lanzhou University, 2011, 47(2): 88-92.
[21] 宁志英, 李玉霖, 杨红玲, 张子谦. 科尔沁沙地优势固沙灌木叶片氮磷化学计量内稳性. 植物生态学报, 2019, 43(1): 46-54. doi: 10.17521/cjpe.2018.0100 NING Z Y, LI Y L, YANG H L, ZHANG Z Q. Nitrogen and phosphorus stoichiometric homoeostasis in leaves of dominant sand-fixing shrubs in Horqin Sandy Land, China. Chinese Journal of Plant Ecology, 2019, 43(1): 46-54. doi: 10.17521/cjpe.2018.0100
[22] FRIEDMAN J. The evolution of annual and perennial plant life histories: ecological correlates and genetic mechanisms. Annual Review of Ecology, Evolution, and Systematics, 2020, 51: 461-481. doi: 10.1146/annurev-ecolsys-110218-024638
[23] 瞿王龙, 杨小鹏, 张存涛, 魏冰. 干旱、半干旱地区天然草原灌木及其肥岛效应研究进展. 草业学报, 2015, 24(4): 201-207. QU W L, YANG X P, ZHANG C T, WEI B. Shrub-mediated “fertile island” effects in arid and semi-arid grassland. Acta Prataculture Sinica, 2015, 24(4): 201-207.
[24] LI Y, ZHOU X, ZHANG Y. Shrub modulates the stoichiometry of moss and soil in desert ecosystems, China. Journal of Arid Land, 2019, 11(4): 579-594. doi: 10.1007/s40333-019-0057-y
[25] NING Z, LI Y, ZHAO X Y, HAN D, ZHANG J. Comparison of leaf and fine root traits between annuals and perennials, implicating the mechanism of species changes in grassland. Frontiers in Plant Science, 2022, 12: 547-778.
[26] 苏强. 浮游动物化学计量学稳态性特征研究进展. 生态学报, 2012, 32(22): 7213-7219. doi: 10.5846/stxb201110141522 SU Q. The framework of stoichiometry homeostasis in zooplankton elemental composition. Acta Ecologica Sinica, 2012, 32(22): 7213-7219. doi: 10.5846/stxb201110141522
[27] 严正兵, 金南瑛, 韩廷申, 方精云. 氮磷施肥对拟南芥叶片碳氮磷化学计量特征的影响. 植物生态学报, 2013, 37(6): 551-557. doi: 10.3724/SP.J.1258.2013.00056 YAN Z B, JIN N Y, HAN T S, FANG J Y. Effects of nitrogen and phosphorus fertilization on leaf carbon, nitrogen and phosphorus stoichiometry of Arabidopsis thaliana. Chinese Journal of Plant Ecology, 2013, 37(6): 551-557. doi: 10.3724/SP.J.1258.2013.00056
[28] EREL R, ANNETTE B, CAPOWIEZ L, DOUSSAN C, ARNAL D, SOUCHE G, GAVALAND A, FRITZ C, ERIC J W, SALVI S V, MARIE L C, HUND A, HINSIGER P. Soil type determines how root and rhizosphere tratis relate to phosphorus acquisition in field-grown mazie genotypes. Plant and Soil, 2017, 412(1): 115-132.
[29] 王彦丽, 范庭, 王旭东, 张育林, 李利敏, 郑洺钧. 长期磷肥不均投入下黄土高原土壤磷素有效性及与土壤性质关系分析. 水土保持学报, 2019, 33(5): 237-242, 250. WANG Y L, FAN T, WANG X D, ZHANG Y L, LI L M, ZHENG M J. Analysis of phosphorus availability and its relationship with soil properties in the Loes Plateau under long-term lmbalanced phosphorus input. Journal of Soil and Water Conservation, 2019, 33(5): 237-242, 250.
[30] 靖淑慧, 刘加珍, 陈永金, 张天举. 滨海湿地不同群落对土壤磷的有效性影响研究. 土壤通报, 2018, 49(2): 392-401. JING S H, LIU J Z, CHEN Y J, ZHANG T J. Effects of plant communities on soil phosphorus availability in coastal wetlands. Chinese Journal of Soil Science, 2018, 49(2): 392-401.
[31] ZHANG K, SU Y, YANG R. Variation of soil organic carbon, nitrogen, and phosphorus stoichiometry and biogeographic factors across the desert ecosystem of Hexi Corridor, northwestern China. Journal of Soils and Sediments, 2019, 19(1): 49-57. doi: 10.1007/s11368-018-2007-2
[32] 孙临泉, 陈子学, 张洪立, 刘冰, 刘丽丽. 微生物植酸酶对土壤有机磷组分含量及有效性的影响. 天津师范大学学报(自然科学版), 2011, 31(2): 86-90. SUN L Q, CHEN Z X, ZHANG H L, LIU B, LIU L L. Effects of microbial phytase on soil organic phosphorus component and its availability. Journal of Tianjin Normal University (Natural Science Edition), 2011, 31(2): 86-90.
[33] RICHARDSON A E, SIMPSON R J. Soil microorganisms mediating phosphorus availability update on microbial phosphorus. Plant Physiology, 2011, 156(3): 989-996. doi: 10.1104/pp.111.175448
-
期刊类型引用(9)
1. 万建琦,丁瑜,任晗,李雯霞,杨勇,黄金盟,关长飞. DdSOC1基因调控德阳柿成花分子机制. 果树学报. 2025(02): 276-287 .  百度学术
百度学术
2. 杨胤延,孙东宇,蔡昕成,胡志群,吴国麟,周碧燕. 莲雾调控开花基因SsSOC1的克隆及表达分析. 热带作物学报. 2025(03): 553-562 . 百度学术
3. 龙丽君,马英姿,曾慧杰,李昌珠,张岗,刘思思,李依民. 灰毡毛忍冬花器官发育相关LmSOC1基因克隆及表达分析. 中南林业科技大学学报. 2024(05): 181-190 . 百度学术
4. 俞亚欣,龙丽君,李昌珠,曾慧杰,乔中全,刘思思,马英姿. 灰毡毛忍冬AP1同源基因的克隆及互作蛋白鉴定. 药学学报. 2024(10): 2880-2888 . 百度学术
5. 梁婷,阙远慧,喻奇伟,贾傛吏,邹广,郭凯阳,黄莺,刘仁祥. 烤烟品种'韭菜坪2号'叶片数的遗传分析及基因定位. 农业生物技术学报. 2023(01): 50-60 . 百度学术
6. 崔秀文,刘迪,黄天苗,李美玲,栗孟飞,魏建和. 当归MADS-box生物信息学及SOC1克隆与表达分析. 中草药. 2023(05): 1551-1560 . 百度学术
7. 蔺悦,陈俊兴,红梅,陈一潇,徐宛玉,张雅茜,包福海,乌云塔娜,王淋,包文泉. 西伯利亚杏PsSOC1-like基因密码子偏好性与进化分析. 内蒙古农业大学学报(自然科学版). 2023(01): 13-25 . 百度学术
8. 刘朝斌,李荣,陈诗婷,张茜,张振源. 核桃开花相关基因JrSOC1的筛选及其在开花中的作用分析. 西北林学院学报. 2023(04): 97-103 . 百度学术
9. 张丽杰,董天一,吴静雯,张萌萌,贾若雪,刘春平. 胡桃楸成花相关基因JmSOC1克隆和表达分析. 沈阳农业大学学报. 2023(06): 673-682 . 百度学术
其他类型引用(11)






计量
- PDF下载量: 0
- 文章访问数: 0
- HTML全文浏览量: 0
- 被引次数: 20

