


{kind=link}
{kind=link}
{kind=link}
CO2浓度升高对不同秋眠型苜蓿叶片解剖结构的影响
[杜乐山 , 刘海燕, 翟晓朦, 任梦云, 李俊生, 关潇]
, 刘海燕, 翟晓朦, 任梦云, 李俊生, 关潇]
, 刘海燕, 翟晓朦, 任梦云, 李俊生, 关潇]
|
|


第一作者:杜乐山(1988-),男,山东高密人,助理研究员,硕士,研究方向为生物多样性保护。E-mail:duleshan@yeah.net
大气CO2浓度升高对植物的影响是目前国内外学者普遍关注的问题。为研究CO2浓度升高对苜蓿( Medicago sativa)不同生长期叶片解剖结构的影响,利用开顶式生长室(open-top chambers,OTC)模拟未来CO2浓度,以秋眠型、半秋眠型和极非秋眠型的苜蓿为试验材料,采用石蜡切片技术研究苜蓿叶片解剖结构的变化。结果表明,1)叶片解剖结构受CO2浓度升高、生长期的影响较大,受秋眠类型影响较小;且受三因子交互效应影响显著。2)叶片解剖结构随CO2浓度升高显著增加( P<0.01),有利于增加叶绿体含量和气体交换速率,进而提高光合作用速率;叶片厚度、表皮厚度、栅栏组织厚度、海绵组织厚度随生长期变化显著增加( P<0.01),但解剖结构在不同秋眠型间差异不大( P>0.05),可能是因为秋眠性受光照时长、气温的影响较大,受CO2的影响较小。
Effect of elevated atmospheric CO2 concentration on plants has aroused wide concern among scholars. In this study, open-top chambers (OTC) were used to model the future CO2 concentration to study the effect of CO2 enrichment on the leaf anatomical structure of three types fall dormancy at different growth stages by using the paraffin section technique. Results showed that: the anatomical structure of leaves was significantly affected by the increase of CO2 concentration, the growth period, and the interaction effect of three factors. The leaf anatomical structure increased significantly with the increase of CO2 concentration ( P<0.01), which was beneficial to increase chloroplast content,gas exchange rate, and subsequently the photosynthesis rate. Leaf thickness, cuticle thickness, palisade tissue thickness, and sponge tissue thickness increased significantly with the growth period ( P<0.01). The anatomical structure of leaves showed no significant differences among three fall dormancy types ( P>0.05), mainly because fall dormancy is influenced by the duration of light and temperature and is less affected by CO2. The results of this study provide a basis for predicting the effect of elevated CO2 on the anatomical structure of plant leaves for adaptation to CO2 concentration.
自工业革命以来, 随着大量伐木毁林以及化石燃料的燃烧, 大气层中CO2浓度日益增高, 已由工业革命前的280 μ mol· mol-1增加到如今的370 μ mol· mol-1, 并以每年1.8 μ mol· mol-1的速度迅速增加[1]。预计到21世纪中期, 大气中的CO2浓度将达到550 μ mol· mol-1[2]; 到21世纪末, CO2浓度将增至421~936 μ mol· mol-1[3]。作为植物光合作用和初级代谢的基本物质, CO2浓度升高是全球气候变化中极少数对农作物生产力有正向因子的作用之一[4], 对植物表型特征、光合作用、生理生化乃至繁殖等均有显著影响。植物叶片是植物进化过程中对环境变化较敏感的器官, 其解剖结构最能体现环境因子对植物的影响[5]及植物对环境的适应策略[6, 7, 8], 因此, 研究CO2浓度升高对植物叶片解剖结构的影响尤为重要。
苜蓿(Medicago sativa)是多年生豆科植物, 具有适应性强、草质优良、营养丰富、适口性好等诸多优点, 被誉为“ 牧草之王” 。秋眠性是苜蓿的重要特性, 北美科学家先后根据秋眠类型将苜蓿划分为9个和11个等级。秋眠型苜蓿秋季刈割后再生速度慢, 产量较低, 抗寒性强; 非秋眠型苜蓿刈割后再生速度快, 产量高, 但越冬率低; 半秋眠型苜蓿介于二者之间[9]。国内外针对不同秋眠类型的苜蓿也开展了大量研究, 特别是翟晓朦等[10, 11, 12, 13]针对CO2浓度升高对不同秋眠类型苜蓿生长及生理的影响开展了一系列研究, 但CO2浓度变化、秋眠类型及生长期对苜蓿叶片解剖结构影响的研究仍十分欠缺。为此, 本研究选取具有代表性的3类秋眠级苜蓿(秋眠型、半秋眠型、极非秋眠型)作为试验材料, 采用开顶式生长室(open-top chambers, OTC)模拟大气CO2浓度增加对苜蓿不同秋眠型、不同生长期叶片解剖结构的影响及苜蓿对CO2浓度变化的响应, 为深入探讨气候变化对植物影响及植物的生态适应性提供数据支撑。
试验所用苜蓿种子由北京林业大学提供, 采用美国标准秋眠性苜蓿对照品种:秋眠型Maverick(简称FD1)、半秋眠型ABI 700(简称FD6)和极非秋眠型UC 21465(简称FD11), 代表3个不同的秋眠类型。
试验在中国环境科学研究院顺义试验站(38° 39'27.97″ N, 104° 04'58.66″ E)进行, 该地属暖温带半湿润气候, 年平均温度11.2 ℃, 年平均降水量为695.4 mm。试验在全透明的OTC中进行, 气室是由高2.4 m、边长1.15 m的玻璃组成的八边形框架结构[12], 保证充足同质的自然光照、土壤(V腐殖质:V黄黏土:V蛭石=1:1:1)及水分条件(0-20 cm土层土壤含水量平均值在40%~45%)。试验模拟当前、21世纪中期和21世纪末的CO2浓度, 设置3个CO2浓度梯度, 探讨不同秋眠型苜蓿在不同生长期叶片解剖结构的变化。试验组全天通入CO2气体, 通过传感器控制OTC内CO2浓度分别为(550± 100) μ mol· mol-1(T1)和(700± 100) μ mol· mol-1(T2); 对照组全天通入自然风, CO2浓度为(350± 50) μ mol· mol-1(CK)。试验共设置12个OTC, 分为3组; 每个OTC放置8个花盆(口径17 cm, 高度12 cm), 花盆的大小、土壤量均保持一致, 每花盆种植10株苜蓿。于5月5日播种开始试验, 分别于分枝期(6月9日)、初花期(7月18日)、盛花期(8月19日)和成熟期(9月28日)取样测定。
取相同叶位、成熟健康的新鲜叶片进行清洗, 用干纱布擦干, 选取0.5 cm× 1 cm的小片, 采样后立即置于FAA固定液中固定24 h以上。石蜡切片按常规方法以酒精脱水-石蜡包埋-切片(厚度8~10 μ m)-番红固绿对染-加拿大树胶封片的步骤进行[14]。对3个CO2处理(CK、T1、T2)、3种秋眠型(FD1、FD6、FD11)、5个生长期(苗期、分枝期、初花期、盛花期、成熟期)各选10张切片, 每一切片选取5个视野, 共获取3× 3× 5× 50=2 250个视野数据。
在OLYMPUS光学显微镜下, 分别测量表皮细胞厚度(epidemiscells thickness, ET)、栅栏组织厚度(palisade parenechyma thickness, PT)、海绵组织厚度(spongy parenchyma thickness, ST)和叶片总厚度(leaf thickness, LT), 并分别计算叶片组织结构紧密度(cell tense ratio, CTR, %)和叶片组织结构疏松度(spongy ratio, SR, %):
CTR=PT/LT× 100%;
SR=ST/LT× 100%。
叶片解剖结构数据均用平均值± 标准差的形式表示。采用Three-way ANOVA分析CO2浓度、秋眠型、生长期及交互效应对叶片解剖结构的影响。分别计算不同CO2浓度、不同秋眠类型、不同生长期的解剖特征平均值, 采用One-way ANOVA分别对CO2处理、秋眠类型和生长期间的差异进行分析, 并利用LSD法进行多重比较。采用One-way ANOVA分析同一秋眠型、同一生长期植株在不同CO2浓度下的差异, 并利用LSD法进行多重比较。数据采用SPSS 19.0软件进行统计分析, 用Origin 9.0软件作图。
苜蓿解剖结构受CO2浓度、生长期的影响较大, 受秋眠类型影响较小(表1)。CO2处理对叶片厚度、表皮厚度、栅栏组织厚度、海绵组织厚度、组织结构紧密度、组织结构疏松度有显著影响(P< 0.01)或极显著影响(P< 0.001); 秋眠类型仅对海绵组织厚度有显著影响(P< 0.05); 不同生长期对所有叶片解剖结构均有极显著影响(P< 0.001)。叶片解剖结构不仅受单因素的影响, 同时也受到多因子共同作用的影响。如CO2处理和生长期的交互效应, 对叶片厚度、栅栏组织厚度、海绵组织厚度有极显著影响(P< 0.001), 对其他指标影响不大; CO2处理与秋眠类型的交互效应对叶片厚度(P< 0.01)、海绵组织厚度有显著影响(P< 0.05), 对其他指标影响不大; 生长期与秋眠类型的交互效应和三者的交互效应对叶片厚度、表皮厚度、栅栏组织厚度、海绵组织厚度、组织结构紧密度有显著(P< 0.01)或极显著影响(P< 0.001), 对SR影响不大。
| 表1 CO2浓度、秋眠型、生长期对叶片解剖结构的影响 Table 1 Effect of CO2 concentration, fall dormancy and growth stage on leaf anatomical structure |
随CO2浓度升高苜蓿叶片平均厚度显著增厚(P< 0.05), T1、T2较CK分别增加了14.9%、22.1%, 且三者间差异均显著(P< 0.05, 表1, 图1)。不同秋眠型叶片平均厚度差异不显著(P> 0.05, 表1)。叶片平均厚度随生长期的延迟显著增厚(P< 0.05), 分枝期、初花期、盛花期、成熟期较苗期分别增加了17.4%、31.5%、53.6%、67.1%, 且不同生长期之间差异显著(P< 0.05, 表1, 图1)。
 | 图1 不同浓度CO2处理对叶片厚度和表皮厚度的影响不同小写字母代表同一秋眠型、同一生长期植株在不同CO2浓度处理之间差异显著(P< 0.05)。下同。Fig. 1 Effect of different CO2 concentrations on leaf and epidermal thicknessDifferent lowercase letters indicate significant difference among different CO2 concentration with the same fall dormancy and growth period; similarly for the following figures. |
随CO2浓度升高苜蓿表皮平均厚度显著增厚(P< 0.05), T1、T2较CK分别增加了4.0%、8.6%, 且三者之间差异均显著(P< 0.05, 表1, 图1)。不同秋眠型表皮平均厚度差异不显著(P> 0.05, 表1)。表皮平均厚度随生长期的变化显著增厚(P< 0.05), 分枝期、初花期、盛花期、成熟期较苗期分别增加了8.3%、14.3%、21.3%、26.3%, 且不同生长期之间差异显著(P< 0.05, 表1, 图1)。
随CO2浓度升高苜蓿栅栏组织平均厚度显著增厚(P< 0.05), T1、T2较CK分别增加了17.6%、25.4%, 且三者之间差异均显著(P< 0.05, 表1, 图2)。不同秋眠型栅栏组织平均厚度差异不显著(P> 0.05, 表1)。栅栏组织平均厚度随生长期的变化显著增厚(P< 0.05), 分枝期、初花期、盛花期、成熟期较苗期分别增加了20.2%、40.1%、64.5%、80.1%, 且不同生长期之间差异显著(P< 0.05, 表1, 图2)。
随CO2浓度升高苜蓿海绵组织平均厚度显著增厚(P< 0.05), T1、T2较CK分别增加了17.8%、25.5%, 且三者之间差异均显著(P< 0.05, 表1, 图2)。不同秋眠型海绵组织平均厚度差异不显著(P> 0.05, 表1)。海绵组织平均厚度随生长期的变化显著增厚(P< 0.05), 分枝期、初花期、盛花期、成熟期较苗期分别增加了19.7%、31.8%、60.4%、76.6%, 且不同生长期之间差异显著(P< 0.05, 表1, 图2)。
 | 图2 不同浓度CO2处理对栅栏和海绵组织厚度的影响Fig. 2 Effect of different CO2 concentrations on palisade and spongy tissue thickness |
随CO2浓度升高组织结构紧密度平均值显著增大(P< 0.05), T1、T2较CK分别增加了1.9%、2.6%, 且T1、T2间差异不显著, 与CK差异显著(P< 0.05, 表1, 图3)。不同秋眠型苜蓿组织结构紧密度平均值差异不显著(P> 0.05, 表1)。组织结构紧密度平均值随生长期的变化差异显著(P< 0.05), 分枝期、初花期、盛花期、成熟期较苗期分别增加了2.1%、6.5%、7.1%、7.7%; 苗期组织结构紧密度最小, 与其他4个生长期差异显著; 成熟期组织结构紧密度最大, 仅与苗期、分枝期差异显著(P< 0.05, 表1, 图3)。
随CO2浓度升高组织结构疏松度平均值显著增大(P< 0.05), T1、T2较CK分别增加了2.23%、2.26%, 且T1、T2间差异不显著, 与CK差异显著(P< 0.05, 表1, 图3)。不同秋眠型组织结构疏松度平均值差异不显著(P> 0.05, 表1)。组织结构疏松度平均值随生长期的变化显著(P< 0.05), 分枝期、初花期、盛花期、成熟期较苗期分别增加了1.6%、0.3%、4.2%、5.5%; 初花期组织结构疏松度均值最小, 成熟期组织结构疏松度最大(P< 0.05, 表1, 图3)。
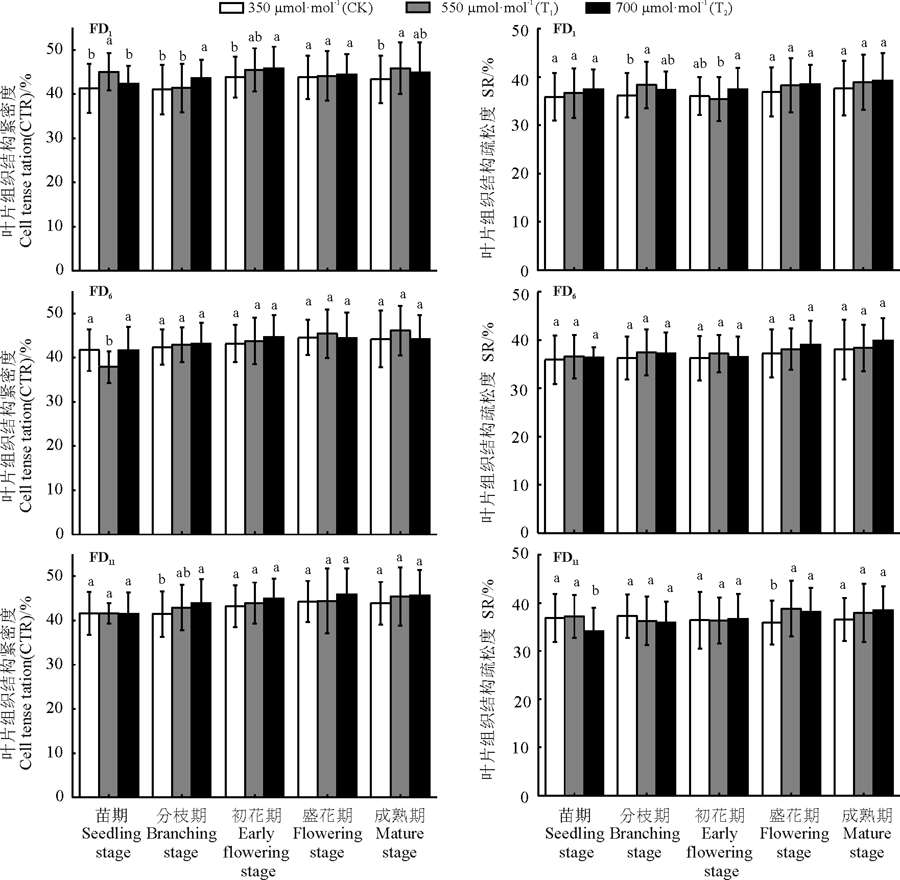 | 图3 不同浓度CO2处理对CTR和SR的影响Fig. 3 Effect of different CO2 concentrations on CTR and SR |
植物叶片解剖结构是研究植物对不同生境的结构与功能适应性的重要材料, 特别是研究植物对大气CO2变化的响应具有重要意义。CO2是植物进行光合作用和初级代谢的基本物质, CO2加富对植物叶片解剖结构有明显影响, 大量研究表明CO2浓度升高显著增加了C3植物叶片厚度、栅栏组织厚度、海绵组织厚度[14], 该结论在本研究得到了进一步证实, 苜蓿叶片总厚度和各组织厚度均有明显加厚。栅栏组织应对CO2加富的方式可分为两种:CO2浓度升高促进植物叶片细胞的分裂, 使栅栏组织层数增加[15, 16]; CO2浓度升高使栅栏组织、海绵组织细胞的体积变大, 叶片厚度增加[17]。栅栏组织层数及厚度是衡量植物光合作用强弱的重要指标[18, 19], 因为栅栏组织含有大量的叶绿体, 其层数及厚度增加能提高植物叶片对光合和环境CO2的吸收能力。海绵组织通常由一些排列较疏松、不规则形细胞组成, 胞间隙发达, 主要进行气体交换[20]。本研究发现, 高CO2浓度下苜蓿的栅栏组织层数未发生变化, 但明显增厚, 海绵组织增厚, 这是苜蓿叶片应对CO2浓度升高的适应性策略[21], 因为栅栏组织和海绵组织加厚, 增加了叶绿体含量和气体交换速率, 进而提高光合作用速率和适合度, 该结论也在翟晓朦等[11]的研究中得到了验证。
秋眠性是苜蓿的重要特性, 由于秋季光照时长变短和气温下降, 其茎枝出现匍匐生长、生长减缓甚至停止。已有研究发现, 光周期[20, 22, 23, 24]、温度[25, 26, 27]等对不同秋眠类型苜蓿有显著影响, 但本研究不同CO2浓度对苜蓿叶片解剖结构的影响不大。可能是因为叶片是对CO2变化最敏感的器官[28], 能够准确反映出CO2对其产生的直接影响; 而秋眠性受光照时长、气温的影响较大, 受CO2的影响相对较小。
不同功能型植物在同一生长期, 叶片解剖结构可能完全不同; 同一植物的不同发育阶段, 叶片解剖结构也会有所差异[29]。如银杏(Ginkgo biloba)在展叶期无栅栏组织和海绵组织的分化; 展叶后叶肉分化为栅栏组织1~2层, 细胞呈长椭球形, 海绵组织发达, 细胞呈横向排列的椭球形; 衰老期部分海绵组织细胞变小, 并纵向排列, 通气系统发达[30]。沙芥(Pugionium cornutum)幼苗期栅栏组织细胞间隙大, 海绵组织细胞排列紧密, 几乎没有细胞间隙; 旺盛生长期海绵组织细胞形状不规则, 具细胞间隙[31]。连香树(Cercidiphyllum japonicum)幼叶发育早期叶肉组织无明显分化, 成熟叶片的栅栏组织细胞在层数和细胞形态上与分化初期差异不大, 但细胞壁较幼期厚, 细胞经向有一定伸长[29]。多年生黑麦草(Lolium perenne)在生长旺盛季, 叶肉细胞的生长与CO2浓度升高呈正相关关系, 而在生长缓慢季, 叶片细胞的生长与CO2浓度呈负相关关系[32]。本研究中, 苜蓿随着时间变化, 其叶片厚度、栅栏组织、海绵组织均有一定程度加厚, 这与其在不同生长发育阶段对生活环境的适应密切关联。因为不同发育阶段的细胞对CO2的传递阻力不同[18], 而叶肉组织通过细胞对CO2的传递阻力来影响叶片的光合作用[33], 苜蓿叶片各解剖结构的协同变化提高了叶片本身的光合能力[10], 进而提高整个植物的生物量[11]。
综上所述, 叶片解剖结构受CO2浓度变化、生长期的影响较大, 受秋眠类型影响较小; 且受三因子交互效应影响显著。解剖结构厚度随CO2浓度升高显著增加(P< 0.01), 这有利于增加叶绿体含量和气体交换速率, 进而提高光合作用速率和牧草产量, 是苜蓿应对CO2浓度增加的适应性反应; 叶片厚度、表皮厚度、栅栏组织厚度、海绵组织厚度随生长期变化显著增加(P< 0.01), 这可能是苜蓿叶片解剖结构协同变化的结果; 但叶片解剖结构在不同秋眠类型间差异不大(P> 0.05), 是因为秋眠性受光照时长、气温的影响较大, 受CO2的影响相对较小。本研究为预测CO2浓度变化对植物的影响提供了重要的数据支撑, 但在自然条件下, 伴随CO2浓度升高, 土壤肥力、水分及土壤微生物等均会发生变化, 研究解剖结构受多个因子的综合影响, 对准确预测未来气候变化对植物的影响及植物的适应策略具有重要意义, 其中涉及的相互作用机理尚待生理和分子方面的进一步研究。
(责任编辑 张瑾)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|