


{kind=link}
{kind=link}
{kind=link}
不同生境角果毛茛种群生物量分配与生长分析
[贾风勤 , 张娜, 纳森巴特]
, 张娜, 纳森巴特]
, 张娜, 纳森巴特]
|
|


于2015年5月采用野外生态学实验方法和室内分析相结合方法对伊犁塔克尔莫乎尔沙漠四爪陆龟保护区和林地生境内短命植物角果毛茛( Ceratocephalus testiculatus)种群生物量分配和生长进行比较研究。结果表明,保护区样地角果毛茛种群株高、总生物量、叶、花葶和果序生物量均显著高于林地( P<0.05),根生物量无差异( P>0.05);但根、叶和花葶生物量分配却显著低于林地( P<0.05),仅果序生物量分配显著高于林地( P<0.05)。两样地内叶、花葶和果序生物量随总生物量增加呈幂函数形式增长,根生物量呈线性增长,均以林地增长速率为高;生殖分配Ⅰ(果序生物量/总生物量)和Ⅱ(花葶生物量/总生物量)所占比值相对稳定,受株高和总生物量变化影响甚小。保护区样地叶生物量变化显著影响着根生物量和生殖生物量( P<0.05),而在林地内仅极显著影响着生殖生物量( P<0.01)。角果毛茛种群通过调节营养构件和生殖构件生物量及其分配速度以适应多变环境,保证在短的生长期内达到生殖成功并使生殖分配比率最大。
The ephemeral species Ceratocephalus testiculatus was mainly distributed in Takeermohuer Desert at Yili in Xinjiang. A nature reserve plot and an artificial woodland were chosen to investigate biomass allocation and growth rhythm of C. testiculatus, based on field investigation and laboratory analysis. The results indicated that height, total biomass, leaf biomass, scape biomass, and frutescence biomass at the nature reserve plot were significantly higher than that in the artificial woodland. However, root, leaf, and scape biomass allocation were significantly lower, and only frutescence biomass allocation was significantly higher. Biomass of leaf, scape, and frutescence increased as a power function with increase in total biomass in each plot. A linear relationship between root biomass and total biomass was observed, and the highest rate of increase was evident in the artificial woodland. The rates of reproductive allocationⅠand Ⅱ (i.e., frutescence biomass/total biomass and scape biomass/total biomass, respectively) were relatively stable, and height and total biomass of the plants hardly affected them. Changes in leaf biomass significantly affected root biomass and reproductive biomass in the nature reserve plot. However, it affected reproductive biomass of plants in the artificial woodland significantly. C. testiculatus populations adjust biomass of vegetative and reproductive modules and allocation rates for adapting to different environments, and ensuring reproductive success and attainment of the largest reproductive allocation ratio in the short span of growth.
植物在有性生殖过程中需要不断地协调营养器官和生殖器官间资源分配以形成最佳资源分配格局, 提高其生存适合度和竞争能力[1, 2]。生物量分配是生殖生态学研究的主要内容, 反映着植物在特定环境条件下的选择适应[3]。生殖分配是指植物一年所同化的物质或固定的能量中用于生殖的比例, 即分配到生殖器官中的有机物重量占整个同化产物的百分比, 作为植物生长特性的一个重要指标, 植物生殖分配格局及其调节生殖分配策略不但受植物生活型差异的影响[4], 也受土壤水分、光照强度等环境因子差异的影响[5]。
短命植物是生活于极端干旱自然环境, 利用冬春雨雪水在春末夏初迅速完成生活周期的一类特殊草本植物, 在我国主要分布在准噶尔荒漠及伊犁河谷地带, 是该区域荒漠中主要的草本植物并成为维持地表稳定、物种多样性和生态系统稳定性的重要贡献者[6, 7]。由于特殊的生存环境, 短命植物通常经过短暂的营养生长后很快进入生殖生长期[8], 短命植物种群生物量分配及生殖再分配情况将会是怎样的?与其生物学特征如株高、总生物量等有何种相关生长关系?
毛茛属短命植物角果毛茛是塔克尔莫乎尔荒漠地区草本植物层片的优势种群, 也是濒危物种四爪陆龟主要取食植物之一[9]。国内近年对毛茛属的研究主要集中于两大方面, 一是在毛茛属植物种质资源、花粉形态、核型分析等系统分类和进化方面有深入研究[10, 11, 12]; 二是对毛茛属植物化学成分、总黄酮含量等药用价值方面的应用研究较多[13, 14], 但关于毛茛属植物生物量及其分配方面的研究很少。本研究以濒危物种四爪陆龟主要取食植物角果毛茛(C. testiculatus)为研究对象, 通过对其生殖构件生物量分配及生殖再分配进行研究, 揭示短命植物营养构件和生殖构件生物量及其分配、生殖生物量再分配对异质生境的响应策略, 同时也为深入了解短命植物的适应策略和生存对策提供科学依据, 并为不同生活型植物生殖分配特征的对比研究提供参考。
研究区位于塔克尔莫乎尔沙漠(80° 27'00″-80° 51'28″ E, 43° 50'34″-44° 09'00″ N)(又名霍城沙漠), 整个沙漠占霍城县土地总面积的9%, 为伊犁河谷面积最大的沙漠, 处于典型大陆性温带荒漠气候带[15]。沙漠地貌形态主要为抛物线状的固定和半固定沙丘组成, 高度通常为3~8 m。年均气温为9.9 ℃, 夏季炎热、干旱, 最高温度达40.2 ℃; 冬季寒冷, 最低温度为-31.3 ℃; 年均降水量254.9 mm, 年均蒸发量约1 510 mm[15, 16]。冬季该区域地表普遍存在10~25 cm积雪, 积雪融化和春季降水使土壤水分含量达到最高保证了荒漠草本植物(包括一年生长营养期、短营养期和多年生草本)在早春能够较好地萌发和生长。调查样地设在沙漠东南部四爪陆龟保护区及保护区外南侧坡底的人工林地。保护区内植物群落类型为银沙槐(Ammodendron argenteum)+心叶驼绒藜(Calligonum densum)群落, 角果毛茛(Ceratocephalus testiculatus)+囊果苔草(Carex physodes)群落等[15]; 人工林地以杨属(Populus)、榆属(Ulmus)树种为主, 林下植物以庭荠(Alyssum desertorum)和角果毛茛为优势种。
2015年5月, 于角果毛茛籽实成熟期, 在其形成的单优种群斑块上做广泛取样。分别从每个样地随机挖取30株, 共计60株, 取样时将植株地上部分连同地下根系一并挖出, 于室内用流水冲洗干净晾干, 注意保持植株完整性; 测定株高后, 将果实、叶、花葶、果序和根剪下单独置于纸袋中并进行编号(来自同一植株的各部分编号相同), 在烘箱内75 ℃烘干至恒重后分别称重; 生物量分配的数量指标根据以下公式计[17, 18, 19, 20]算。
生产分配(productive allocation, PA):PA=BL/BT× 100%;
根生物量分配(root biomass allocation, RA):RA=BR/BT× 100%;
生殖分配(reproductive allocation, REA):
REA=(BF+BSC)/BT× 100%;
生殖分配Ⅰ (reproductive allocation Ⅰ , REAⅠ ):REAⅠ =BF/BT× 100%;
生殖分配Ⅱ (reproductive allocation Ⅱ , REAⅡ ):REAⅡ =BSC/BT× 100%.
式中:BL(leaf biomss)为叶生物量; BR(root biomass)为根生物量; BSC (scape biomass)为花葶生物量; BF(frutescence biomass)为果序生物量; BT(total biomass)为总生物量。
对植株个体构件生物量与总生物量、生殖构件生物量分配与株高和总生物量间关系是在拟合的线性、指数和幂3种函数中, 选用相关性最高的作为关系模型[18]。
用SPSS 17.0对根、叶、花葶、果序、株高及生殖生物量分配进行相关性分析。采用单因素方差分析(One-way ANOVA)比较样地间茎、叶、花葶和果序生物量的差异显著性, 变异系数(CV)反映样本的相对变异度[18]; 用Excel软件绘图。
测定结果显示, 角果毛茛种群株高在两个样地间差异显著, 林地种群植株较保护区高大且整齐, 林地株高为(6.57± 0.93) cm, 变异系数为14.1%, 较保护区种群平均高出1.47 cm(P< 0.05), 变异系数则低5.6%。单株总生物量在两样地间也差异显著, 保护区角果毛茛种群的单株生物量较林地种群平均高106.3 g(P< 0.05), 变异系数也高出8.85%, 如果把变异系数和观测值的范围作为表型可塑性的数量指标, 则保护区样地种群的株高和生物量的表型可塑性普遍大于林地生境。进一步统计分析表明, 构成总生物量的叶、花葶和果序生物量的绝对数量指标在两样地间差异显著(除根生物量外), 均以保护区种群为高, 其中又以果序生物量为最大, 是林地的2.78倍。从相对数量指标来看, 两样地内角果毛茛种群物质分配策略截然不同, 保护区种群仅果序生物量分配显著高于林地, 其余构件生物量分配均小于林地; 其中又以根生物量分配为最高, 为保护区的1.94倍; 其次为生产分配(叶生物量分配), 是保护区的1.53倍; 最小为花葶生物量, 为保护区的1.35倍(图1)。
 | 图1 角果毛茛种群株高、生物量及其分配分布状况 注:对于每一测定性状, 测定值标注不同字母时表示在两样地间存在显著差异(P< 0.05)。Fig. 1 Structure of biomass, biomass allocation and plant height on C. stesticulatus Note: For each quantitative character, different lowercase letters indicate significantly different between two habitats at the 0.05 level. |
角果毛茛种群营养构件包括叶和根, 两样地叶生物量随总生物量增加呈幂函数形式增长, 从拟合方程参数可知, 随植株生物量增加, 叶生物量增加速率以林地为大(b林地=1.026 3), 是保护区的1.10倍; 根生物量随总生物量增加呈线性增长, 在保护区和林地总生物量每增加1 g, 根生物量分别增加0.011和0.015 g, 林地生物量增加幅度略高于保护区(图2)。角果毛茛种群生殖构件由花葶和果序组成, 两样地花葶和果序生物量随总生物量增加呈幂函数增长, 又均以林地增长速率为快, 分别为保护区的1.27和1.94倍。由此说明, 无论是叶、花葶和果序生物量随总生物量的幂函数增长还是根所表现的线形增长, 林地种群植株构件单位生物量随单位总生物量的增加速率均高于保护区, 表明林地生境种群更侧重于伸长生长以提高其生存能力, 保护区种群则侧重生物量积累以提高其现实竞争力。
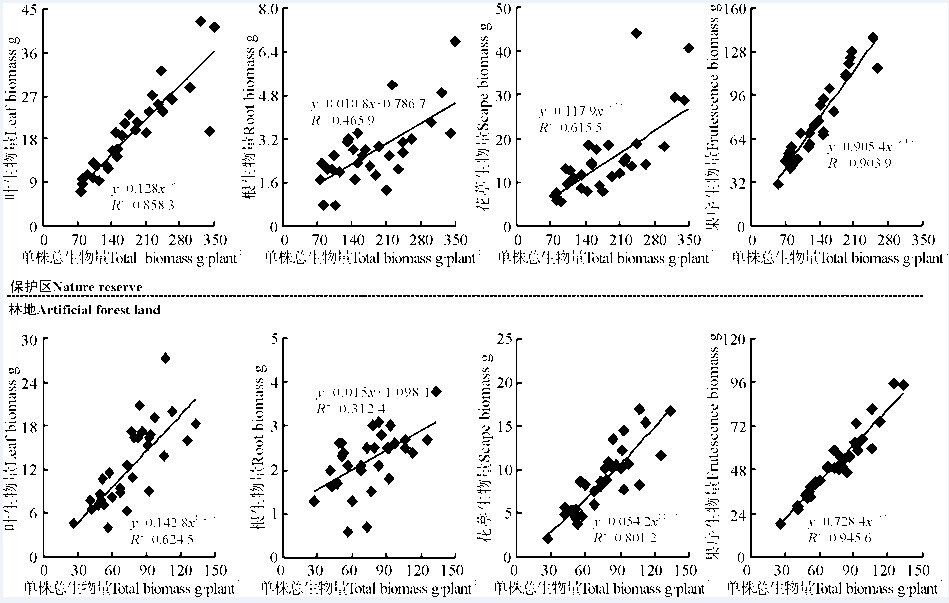 | 图2 不同生境角果毛茛种群构件生物量与植株生物量的观察值及拟合曲线Fig. 2 Relationship between biomass of modules and total biomass of C. testiculatus populations under different habitats |
统计分析显示, 两样地角果毛茛种群生殖分配Ⅰ (花葶)与株高均呈线性正相关关系, 且增加量相近(a保护区=1.591 7≈ a林地=1.568 9); 生殖分配Ⅱ (果序)均呈线性负相关关系, 且种群株高每增加1 cm, 保护区种群生殖分配Ⅱ 减少量为1.474 5, 林地为2.282 2; 生殖生物量分配均呈幂函数正相关增长, 但增长幅度很小约为0(b保护区=0.008 9, b林地=0.061 3)(图3), 由此可知, 在两样地内, 株高的增加均以减少生殖分配Ⅱ 为代价, 林地生境需要消耗的更多, 主要是林地种群株高显著高于保护区, 但无论生殖分配Ⅰ 和Ⅱ 随株高增加或减少, 但生殖分配相对稳定。进一步分析表明, 两样地角果毛茛种群生殖分配Ⅰ 和Ⅱ 与总生物量也呈幂函数或线形函数的正相关或负相关关系, 总体上总生物量的增加对生殖分配(a保护区=0.004 9, a林地=0.018 2)、生殖分配Ⅰ (b保护区=-0.074 5, b林地=0.154 1)和Ⅱ (a保护区=0.005 8, b林地=-0.009 4)的影响非常微小, 从生物量分配策略上分析, 角果毛茛将大部分生物量分配给生殖生长, 且分配比例相当稳定, 只有较少生物量用于营养生长(约20%), 这与多数植物采取的生物量分配策略相一致[13, 14], 也验证了短命植物将投入更多的生物量进行种子生产以克服在荒漠生境中生活史不同阶段可能带来的死亡风险, 使种群得以延续和扩大。
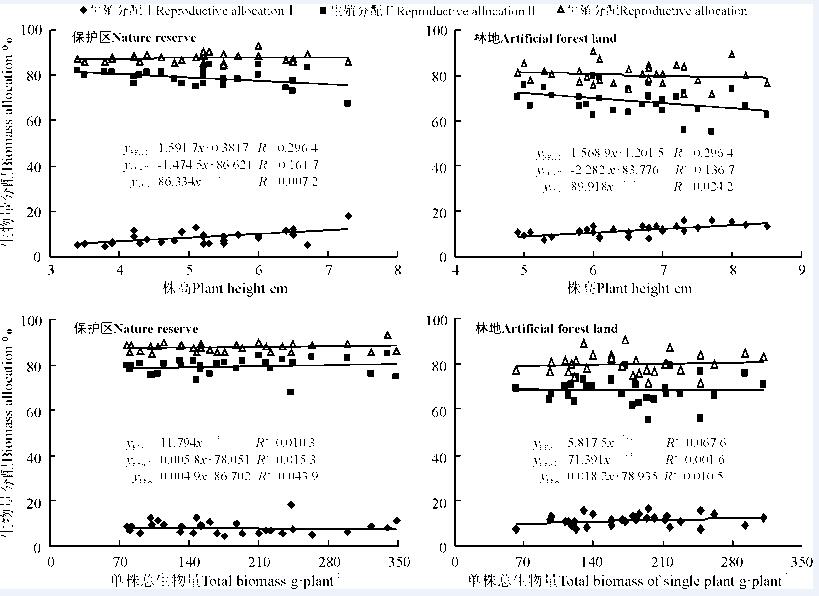 | 图3 不同样地内角果毛茛生殖生物量分配与总生物量间拟合曲线Fig. 3 The curve fitting of relationship between reproductive biomass allocation and total biomass of C. testiculatus populations in different habitats |
植物生长过程中, 各构件之间都存在相互联系、相互制约的关系。对各样地角果毛茛种群根生物量、叶生物量和生殖生物量间进行简单线形相关分析, 无论保护区样地还是林地样地, 3个变量间均存在极显著正相关关系(P< 0.01)。为了进一步阐明消除某一变量取值对变量间两两关系的影响, 在变量间进行偏相关分析, 可知控制生殖生物量的影响后, 根和叶生物量的相关关系在保护区为显著相关(P< 0.05), 在林地内则不存在显著相关关系(P> 0.05); 控制根生物量、叶生物量与生殖生物量在两样地内均呈极显著正相关关系; 控制叶生物量, 根生物量与繁殖生物量在两样地内均不存在显著相关关系。表明在保护区样地叶生物量变化显著影响着根生物量和生殖生物量, 而在林地内极显著影响着生殖生物量(表1)。
| 表1 角果毛茛茎、叶和繁殖生物量3个变量的简单相关系数和偏相关系数 Table 1 Simple and partial correlation coefficient of stem, leaf, and reproductive biomass in C. testiculatus |
通常, 当植物受到光照条件限制时, 植物减少对根生物量的分配, 将更多的资源分配到地上部分, 增加茎、叶生物量的分配来获得更多的光照资源, 提高光竞争能力和生存适合度, 例如木荷(Schima superba)幼苗在林窗边缘生长时采取将生物量相对多地分配给叶片以适应光照不足的生境; 而在林窗中央生长的幼苗则将生物量较多地分配给茎, 以实现和维持有利生境下快速生长的需求[21, 22]; 入侵草本植物兰花菊三七(Gynura sp.)[5]和紫南幼苗(Phoebe sheareri)[23]也会在强光生境中减少根和叶生物量分配以增加茎生物量分配。也有相反的研究结果, 如高光照能显著促进胡杨(Popular euphratica)幼苗根的伸长和生物量的积累, 而遮阴则抑制这些变化[24]。还有研究表明, 无论在林窗生境还是在林下生境, 植物地上和地下生物量分配比例无显著差异[25]。这些研究表明, 不同植物对光环境变化所采用的生物量分配策略不是单一的。本研究中, 光照充足的保护区样地角果毛茛种群叶、花葶和果序的生物量均显著高于林地生境, 根生物量无显著差异, 这与胡杨幼苗对光照的响应[24]有所不同。同时, 较绝对数量指标而言, 相对数量指标是生物体间相对生长性能比较中更为有效的定量指标, 因此, 在本研究中尽管保护区植物个体构件生物量均显著高于林地(除根生物量), 但经过对生物量分配进行差异显著性比较发现, 林地内植株根、叶和花葶生物量分配均显著高于保护区, 这与大耳叶风毛菊(Saussurea macrota)、甘西鼠尾草(Salvia przewalskii)和千里光(Senecio scandens)3种植物生物量分配的研究结果[26]相符, 表明为了适应光照强度弱的不利生境, 植物体采取减少果序生物量分配, 增加营养构件生物量分配, 使得其具备较大的竞争力[22]以获取更多的光照资源。
植物生长、繁殖等各种功能的维持过程, 也是对有限资源协调分配的过程, 植物通过调节生长、生殖以及生殖再分配之间的资源分配比率来提高适合度以适应环境中资源的变化[27, 28, 29]。植物种群或个体的生殖分配是植物长期与环境相互作用、进化、选择和适应所形成的, 一方面受植物本身生物学特性所决定如植物生活型、遗传性等, 通常较稳定[30], 同时也会因生境条件变化出现明显的可塑性, 如种群密度、光照强度和土壤水分等[31, 32]。当植物生长分配主要受遗传决定时, 植物生长条件几乎对其无影响, 研究表明, 尽管欧洲千里光(Senecio vulgaris)生长的营养条件不同, 植株生物量的变异系数也达到270%, 但生殖分配却相当稳定, 变化幅度小于2%[33]。生境变化对植物生殖分配的作用也不可忽视, 它反映着植物在环境适应能力上所具有的特性, 对小画眉草(Eragrostis poaeoides)生殖分配研究发现, 在沙地恢复演替过程中, 演替后期的稳定生境有利于植物进行生殖生长, 与演替初期相比生殖分配呈极显著增长[34]。本研究中, 同一生境内角果毛茛种群株高和总生物量的变化对生殖分配及其再分配影响甚小, 由此推断角果毛茛生殖分配主要是由其遗传性所决定; 不同生境中角果毛茛种群生殖分配存在显著差异, 良好生境条件有利于植物生殖分配增加, 由此表明角果毛茛种群生殖分配受其自身遗传性和环境条件的共同作用。
综上所述, 角果毛茛种群通过根、叶、花葶和果序生物量等数量特征的变异以适应保护区和林地生境内光照、水分不均衡。在两生境角果毛茛种群均超过80%的繁殖构件生物量分配进一步验证了短命植物将投入更多的生物量进行种子或附属结构生产以克服在荒漠生境中生活史不同阶段可能带来的死亡风险, 使种群得以延续和扩大[35]。角果毛茛种群物质分配的异速生长规律集中体现了随机环境因子和遗传因子与植物生长调控和物质分配间的协调策略, 这对塔克尔莫乎尔沙漠及其周边环境植被保护和维持具有一定意义。
(责任编辑 武艳培)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|