




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
抗草甘膦转基因玉米AG16分子特征和抗性鉴定
[刘苗苗1 , 程家慧1 , 林海燕1 , 沈志成1 , 王志勇2 , 林朝阳1  ]
]
]
|
|


第一作者:刘苗苗(1990-),女,江西余干人,在读硕士生,研究方向为植物保护与农业转基因技术。 E-mail:21416110@zju.edu.cn
抗草甘膦转基因玉米( Zea mays)能有效降低杂草防治成本,具有重要的应用前景。浙江大学转基因抗虫植物和生物安全实验室前期通过农杆菌介导法,以玉米Hi-Ⅱ品种为受体导入新型抗草甘膦基因 G10evo,获得不同的抗草甘膦玉米转化系。在此基础上,筛选了具有良好草甘膦抗性的转基因玉米转化系AG16。本研究利用PCR、Western杂交、Southern 杂交、ELISA等方法对AG16进行分子特征检测,并对AG16的草甘膦抗性水平进行鉴定。结果表明, G10evo在AG16中为单拷贝插入;G10evo蛋白在AG16的根、茎和叶组织中表达;ELISA分析表明,草甘膦喷洒前,嫩茎中G10evo蛋白的表达量达到9.975 μg·g-1。温室中草甘膦抗性测定结果显示,AG16能抗4~8倍田间浓度的草甘膦,远高于草甘膦在实际生产中的使用量。因此,AG16草甘膦抗性水平达到生产需求,具有产业应用潜力,为培育具有自主知识产权的抗草甘膦玉米提供了种质资源。
Glyphosate-resistant transgenic maize can significantly improve the efficiency of weed control. In previous studies, the newly glyphosate-tolerant gene G10evo was integrated into the genome of maize named Hi-Ⅱ by Agrobacterium-mediated transformation, and different glyphosate-tolerant maize events were obtained. In the study, transgenic maize AG16 was selected for molecular characterization by PCR, Western blot, Southern blot and ELISA. Southern blot analysis using probe for G10evo showed that T-DNA was integrated in a single site in the maize genome. Western blot analysis showed that G10evo protein was expressed in leaf, stem and root of AG16. ELISA assay showed that the expression level of G10evo protein in the young stem reached 9.975 μg·g-1 before glyphosate spraying. Glyphosate spray assay in the greenhouse showed that the AG16 is tolerant to glyphosate at concentrations up to 4 times of recommended field application level. Therefore, the transgenic maize AG16 has the potential of field application, which is expected to become completely independent research and independent intellectual property rights of domestic glyphosate-resistant corn.
玉米(Zea mays)是全球主要的粮食作物之一, 也是世界上产量最高的粮食作物之一[1], 同时, 玉米生产过程中投入少, 产出高, 光合产物总量有50%~60%贮存于秸秆当中, 是理想的饲料和工业原料[2, 3]。自1990年第1株转基因玉米问世以来, 转基因技术在玉米育种中发挥了巨大作用, 定向改良抗病虫、抗逆性、抗除草剂、雄性不育等性状, 在玉米遗传改良方面展示出了良好的应用前景[4]。目前, 玉米是世界上第二大规模的商业化转基因作物, 2016年全球转基因作物种植面积达到1.85亿hm2, 转基因玉米的种植面积达到0.606亿hm2, 占转基因作物总面积的33%[5], 其中转基因抗除草剂玉米发展最迅速, 种植面积最广泛, 2014年抗除草剂转基因玉米的经济效益累计90.5亿美元, 为全球农业生产带来了巨大的经济效益[6, 7]。
草甘膦是农业上使用量最多的除草剂[8], 其有效化学成分为N-磷酸甲基甘氨酸[N-(phosphonomethyl)-glycine, H2O3P-CH2-NH-CH2-CO2H]。1974年首次在美国批准使用, 商品名为Roundup, 目前已在超过100种农作物和近130个国家用于杂草防治[7], 草甘膦在植物中的毒性作用是通过抑制5-烯醇丙酮莽草酸-3-磷酸酯合成酶(EPSPS)实现的, 该途径仅在植物、细菌、真菌和一些寄生虫中发现, 对多种杂草具有高度活性, 但对哺乳动物几乎没有影响, 是一种环境友好型除草剂, 具有低成本、易降解的特性[9]。因此, 草甘膦仍然是应用40多年后使用最广泛的除草剂。草甘膦是高效的灭生性除草剂, 不能用于常规植物的选择性除草, 只能用于抗草甘膦转基因植物的田间除草, 具有时间和空间的限制。因为草甘膦的高除草效率和低成本优势, 抗草甘膦转基因玉米和大豆已经推广二十多年。抗草甘膦性状除了在农作物上具有重要价值, 在草坪草上的价值也越来越得到重视。草坪杂草防治是草坪养护的主要任务, 常规草坪草一般使用选择性除草剂进行除草, 但是选择性除草剂往往难以杀灭与栽培品种近缘的杂草, 发展抗草甘膦转基因草坪草, 可以提高草坪杂草防治效率, 降低草坪维护成本。近些年, 已经有国外生物技术企业研发和发展了抗草甘膦草坪草, 如美国研发的抗草甘膦早熟禾(Poa pratensis)已经具备推广应用条件[10]。
虽然从微生物和杂草中克隆出许多抗草甘膦EPSPS基因, 但截至现在只有CP4 EPSPS, 玉米EPSPS突变体和GRG23突变体应用于商业化推广的转基因作物中, 并被国际农业生物技术应用服务组织(ISAAA)数据库批准[11, 12, 13, 14, 15, 16, 17, 18]。世界上商业化推广最多的抗草甘膦基因主要是CP4 EPSPS基因, 该基因在多种作物上得到广泛的应用。含两个抗草甘膦CP4 EPSPS基因表达框的NK603转基因玉米在2016年已经获得26个国家及欧盟 28 国的54个批文[5]。G10evo是浙江大学转基因抗虫植物和生物安全实验室自主克隆和改良后的抗草甘膦基因, 具有对草甘膦的高度抗性, 可同时作为抗草甘膦性状和转基因筛选标记应用于转基因作物、牧草及草坪草等的研发中, 是极有潜力投入应用的抗草甘膦基因。中国是玉米需求大国, 2015年进口玉米达330万t[19]。目前我国转基因玉米尚未获得商业化生产种植, 发展转基因玉米有利于提高我国玉米自主生产能力, 尤其是发展具有我国自主知识产权的转基因抗草甘膦玉米, 对杂草防治, 节约劳动成本, 提高农民收益具有重要意义。
本研究中的抗草甘膦EPSPS基因G10evo是浙江大学转基因抗虫植物和生物安全实验室具有自主知识产权, 来源于细菌Deinococcus radiodurans R1, 它与目前划分出的EPSPS家族Ⅰ 、Ⅱ 类抗草甘膦基因亲缘性很低, 保守序列相差很大, 既不属于Class Ⅰ 也不属于Class Ⅱ [20, 21]。该基因作为大豆转化体系中的筛选基因和培养抗草甘膦大豆都具有较好的效果[22, 23]。本研究通过抗性鉴定、分子生物学特征分析, 明确抗草甘膦玉米AG16的分子特征和草甘膦抗性水平, 以期为培育自主知识产权的转基因抗草甘膦玉米提供种质资源。
本研究使用的玉米转化受体为Hi-Ⅱ 品种, 转育用亲本自交系为郑 58(Z58)和昌7-2, 种子由浙江大学转基因抗虫植物和生物安全实验室保存并种植, 对照组郑单958是Z58和昌7-2杂交获得的杂交种。转化事件AG16 T0代与Z58杂交后, 以Z58为轮回亲本, 回交4代, 再自交2代, 得到含有草甘膦抗性基因的自交系Z58-AG16, Z58-AG16与昌7-2杂交获得的F1代杂交种用于本研究。
采用PCR扩增方法检测转基因玉米AG16目的基因。采集5个Z58-AG16与昌7-2杂交获得的F1代玉米叶片, CTAB法[24]提取玉米基因组, 用NanoDrop2000 (Thermo Fisher Scientific, Wilmington, DE, USA)检测所提取玉米基因组的质量和浓度。根据目的基因设计引物G10evo-F(GACGCCCTGCCCGCCACCTTC)和G10evo-R(GGCGGTGGCCTCAGCGTACTCG), 扩增产物用含有1% EB的琼脂糖凝胶电泳检测PCR结果, 用凝胶成像系统观察结果并拍照。
Southern blot[25]分析使用Roche的DIG DNA Labeling and Detection Kit Ⅱ 试剂盒(Roche, Basel, Switzerland)。CTAB法提取玉米基因组, 选择适当的两种酶分别酶切100 μ g玉米基因组, 酶切片段用0.8%的琼脂糖凝胶分离, 分离后转带正电的尼龙膜(Amersham, UK), 转膜20 h左右。利用PCR方法扩增合成杂交探针, 扩增的目的片段使用DIG-High Prime DNA Labeling标记。转膜结束后120 ℃烘烤尼龙膜, 30 min后, 将尼龙膜置于杂交管中, 42 ℃预杂交30 min, 倾尽预杂交液, 加入含地高辛标记探针的杂交液, 55 ℃杂交过夜。最后洗膜, 化学曝光显影20 min。
采用Western blot[26]的方法来检测转基因玉米AG16的G10evo蛋白表达情况。采集玉米新鲜叶片0.1 g, 液氮浸泡, 研磨。磨碎的粉末中加入200 μ L PBS, 混匀离心, 取上清, 按体积比加入5× SDS-PAGE Loading Buffer。制备好的蛋白样品用SDS-PAGE电泳分离, 分离后的蛋白转移到硝酸纤维素膜上, 再将膜封闭在含5%的脱脂奶粉的TBST溶液中, 37 ℃摇床1 h。封闭完成后, 换1%的脱脂奶粉的TBST溶液, 向溶液中按1∶ 2 500的比例加入一抗, 37 ℃摇床孵育1 h, 再用TBST洗膜3次, 每次5 min。一抗孵育后在1%的脱脂奶粉的TBST溶液中, 按一定比例加入二抗, 37 ℃摇床孵育1 h。最后洗膜, DAB显色, 显色至出现条带时用水清洗膜, 晾干, 拍照。
在温室种植转基因玉米AG16和非转基因玉米郑单958采用盆栽的方式, 盆的大小为直径(d)30 cm× 高(H)50 cm, 每盆4株, 后期间苗, 每盆保留两株。每盆做好标签, 等到玉米生长到4-5叶期的时候, 分别对其喷施不同浓度的草甘膦, 每个浓度4个重复, 一周后拍照记录。本研究喷洒的草甘膦为美国孟山都公司生产的农达(20150425, Monsanto, LaConner, USA), 是灭生性内吸传导型茎叶除草剂, 通过植物绿色部位吸收。原产地马来西亚, 水剂型, 有效成分含量30%, 草甘膦异丙胺盐含量41%, 玉米田间推荐使用药量为每亩150~250 mL, 每亩合(666.67 m2)兑水30~40 L, 约2.05 g· L-1, 45 mL· m-2。将水剂农达用水按体积比稀释成有效质量浓度为0、2.05 (1∶ 200)、4.10 (1∶ 100), 8.20 (1∶ 50)和16.40 g· L-1 (1∶ 25)共5个梯度, 分别喷洒在转基因玉米和非转基因玉米上, 每盆样品的喷洒量为45 mL·
用ELISA试剂盒(YouLong Biotech, Shanghai)检测转基因玉米不同组织的G10evo蛋白表达情况。操作方法参考G10-EPSPS试剂盒使用手册。从转基因玉米AG16和非转基因玉米郑单958提取根、茎和叶片中的蛋白, 叶片从新叶到老叶取4片叶片(Leaf 1, Leaf 2, Leaf 3, Leaf 4)分别提取蛋白, 每个组织6个重复, 在450 nm下测其吸光度[26]。
任意采集5株转基因玉米植株AG16和1株非转基因郑单958的叶片, 用于PCR鉴定。用CTAB法提取转基因玉米和非转基因玉米基因组, 对载体1300-p35S-pUbi-G10evo(图1)中的T-DNA中的目的基因G10evo进行鉴定, 确定是否含有T-DNA。预计扩增G10evo基因目的条带大小为1 311 bp, 用含有 1% EB的琼脂糖凝胶电泳检测PCR结果。结果显示, 5株AG16转基因玉米基因组中扩增出G10evo基因的目的片段均为1 300 bp左右, 与阳性质粒扩增出的条带大小一致, 非转基因对照组没有出现条带, 这说明5个F1代杂交后代中均含有G10evo基因(图2)。
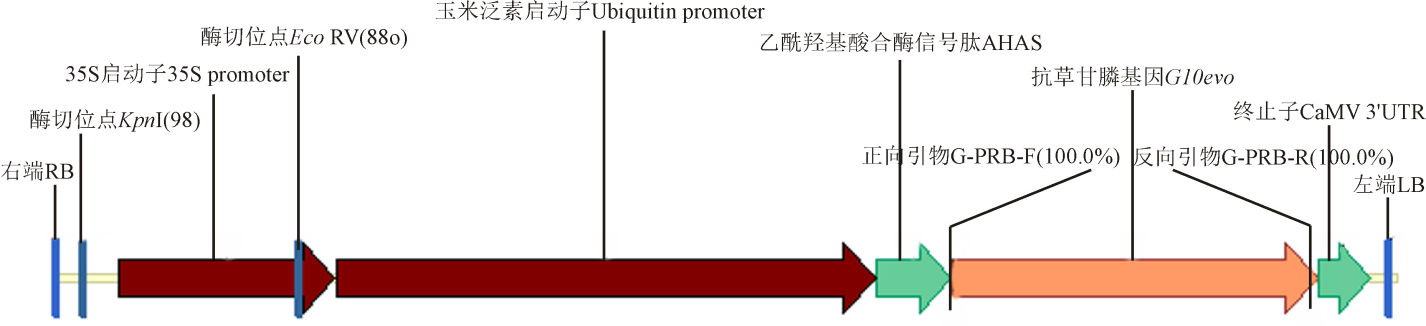 | 图1 载体1300-p35S-pUbi-G10evo图谱Fig. 1 Map of the vector 1300-p35S-pUbi-G10evo |
 | 图2 转基因玉米G10evo PCR鉴定 注:M, 基因Marker; +, 阳性质粒对照; -, 非转基因玉米对照; 1-5, 转基因玉米随机挑选的植株。Fig. 2 PCR analysis of G10evo in GM-maize Note: M, DNA maker (Tran2K plus, Transgen); +, Positive plasmid(CK); -, Non-GM-maize (CK); 1-5, Different GM-maize plants. |
用草甘膦筛选过的转基因阳性玉米, 进行DNA印记检测(Southern blot), 以检测T-DNA的拷贝数。根据G10evo基因序列设计合成探针的引物G10evo-PRB-F(GACGCCCTGCCCGCCACCTTC)和G10evo-PRB-R(GGCGGTGGCCTCAGCGTACTCG)。AG16基因组和玉米阴性对照分别用KpnⅠ 和EcoRⅤ 单酶切, 阳性质粒用KpnⅠ 单酶切, 杂交温度根据GC含量设定为55.5 ℃。阳性质粒酶切片段大小4 536 bp, EcoRⅤ 的酶切片段大小为5 000 bp左右, KpnⅠ 的酶切片段大小为7 400 bp左右, 均符合预期的大小。而且AG16的两种酶切片段的Southern blot结果显示G10evo在玉米基因组中为单拷贝(图3)。
转基因玉米AG16中G10evo的Southern blot分析 Southern blot of G10evo in AG16
注:+, 阳性质粒KpnⅠ 酶切; 1, AG16基因组EcoRⅤ 酶切; 2, 非转基因玉米基因组EcoRⅤ 酶切; 3, AG16基因组KpnⅠ 酶切; 4, 非转基因玉米基因组KpnⅠ 酶切。
Note: +, KpnⅠ digest Positive plasmid; 1, EcoRⅤ digest AG16 genome; 2, EcoRⅤ digest Non-GM Maize genome; 3, KpnⅠ digest AG16 genome; 4, KpnⅠ digest Non-GM Maize genome.
用Western blot方法对抗草甘膦玉米转化系AG16不同组织G10evo蛋白的表达分析结果显示, 抗草甘膦蛋白G10evo在根、茎和叶等组织中均有表达, 目的蛋白分子与原核表达的G10evo大小一致, 为48 kDa, 且叶片和茎中G10evo的表达量明显高于根部(图4)。
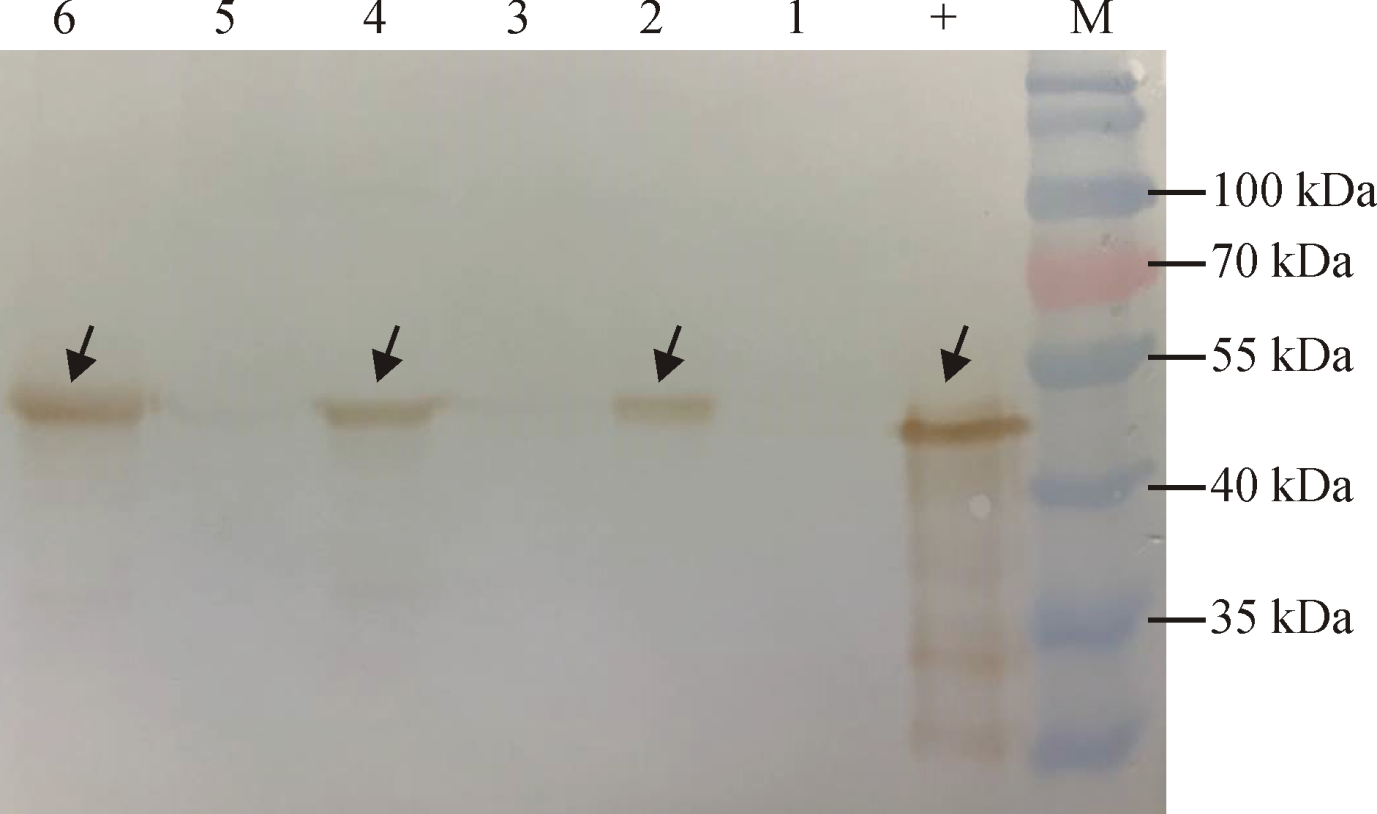 | 图4 Western blot分析检测转基因玉米G10evo蛋白表达 注:M, 预染蛋白Marker; +, 原核表达的G10evo蛋白; 1、3、5, 非转基因玉米的根、茎、叶; 2、4、6, 转基因玉米AG16根、茎、叶。Fig. 4 Western blot analysis of G10evo protein expression in GM-maize Note: M, Prestained protein ladder; +, G10evo protein expressed in E. coli; 1, 3, and 5, Root, stem and leaves of Non-GM-maize; 2, 4, and 6, Root, stem and leaves of GM-maize AG16. |
对生长在温室条件下, 4-5叶期的转基因玉米和非转基因对照玉米分别喷洒相当于大田推荐使用浓度的0、1、2、4和8倍的0、2.05、4.10、8.20和16.40 g· L-1 5个浓度的草甘膦, 一周后观察其表型差异。
在喷施2.05、4.10、8.20和16.40 g· L-1浓度的草甘膦1周后, 非转基因玉米对照组全部死亡; 在喷洒2.05、4.10和8.20 g· L-1浓度的草甘膦1周后转基因玉米, 没有明显的药害(图5), 这说明转G10evo基因玉米具有较高的草甘膦抗性。喷洒16.40 g· L-1浓度的草甘膦对转基因玉米会产生一定的药害, 叶缘和叶尖枯黄、玉米生长缓慢, 个别心叶有白斑, 但持续2~3周后, 药害逐渐消除, 转基因玉米能恢复正常生长。8倍浓度的草甘膦喷施2~3 d后, 转基因玉米叶片上出现轻微的白斑, 这可能是因为草甘膦的毒害作用, 也有可能是因为8倍田间浓度的草甘膦中, 异丙酰胺盐浓度过高, 使得转基因玉米叶片因为渗透压的作用受到伤害, 导致叶片出现轻微白斑。
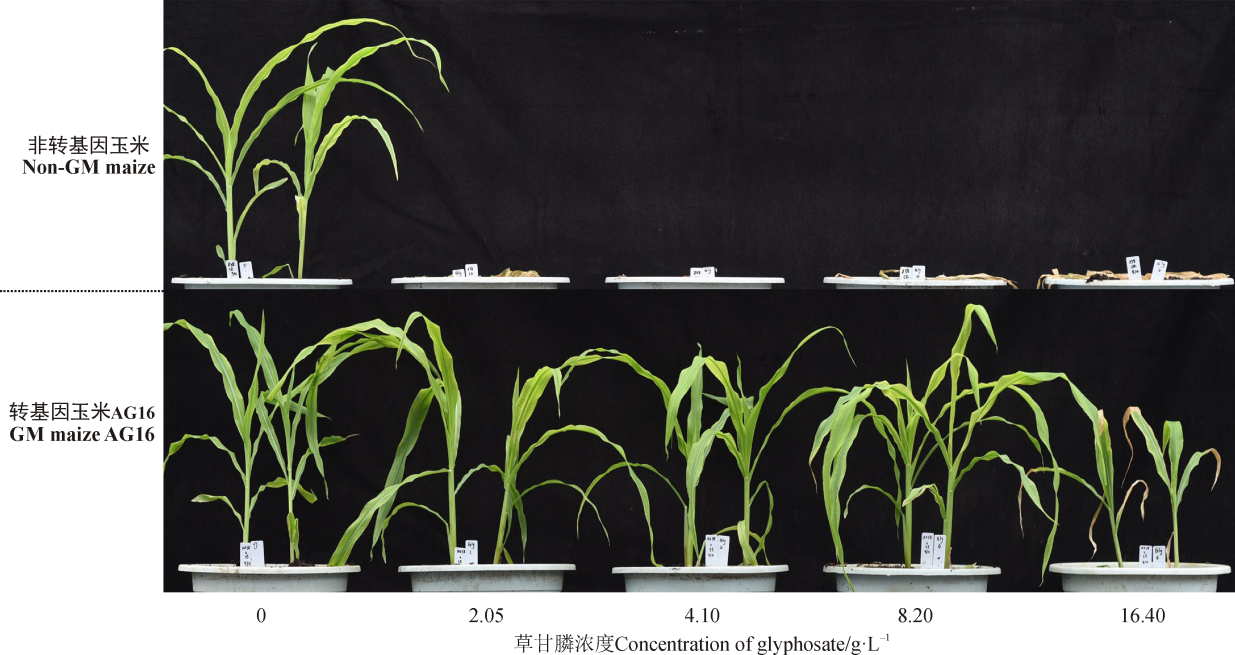 | 图5 转基因玉米AG16的草甘膦抗性Fig. 5 Glyphosate tolerance of GM AG16 maize |
 | 图6 草甘膦喷洒前后AG16玉米不同组织的G10evo蛋白表达量Fig. 6 Concentration of G10evo in different tissues of AG16 maize before and after spraying glyphosate |
对生长在温室条件下, 4-5叶期的转基因玉米AG16不同组织中G10evo蛋白含量进行ELISA定量检测。结果显示, 在喷洒草甘膦前幼茎组织中G10evo蛋白表达量最高, 为9.975 μ g· g-1, 其次是第4片叶, 为8.750 μ g· g-1; 表达量最低的是新生第1和第2片叶, 分别为2.152和2.163 μ g· g-1。喷洒草甘膦后各组织中的G10evo蛋白表达量均有提高, 但表达量最高的还是在幼茎组织中, 为11.234 μ g· g-1, 增加12.6%, 最少的为第1片叶, 为3.315 μ g· g-1, 增加54.0%, 第2片叶为3.524 μ g· g-1, 增加62.9%, 其余增加量均小于10%(图6)。这说明在草甘膦的刺激下, 幼嫩组织能增加G10evo蛋白的表达量, 以提高对草甘膦的耐受能力。
根据氨基酸保守序列和对草甘膦的敏感性, 天然EPSPS被分为两类, 通常来自大多数植物和革兰氏阴性细菌的Ⅰ 类EPSPS被认为是对草甘膦敏感, 但是由于某些氨基酸突变, 它们因此对草甘膦产生耐受性[27]。 相比之下, 来自根癌土壤杆菌的CP4菌株, 无色杆菌LBAA菌株, 假单胞菌属 PG2982菌株, 枯草芽孢杆菌, 金黄色葡萄球菌和奥氏嗜热盐丝菌 H168本身具有对草甘膦的耐受性, 与Ⅰ 类EPSPS相比同源性低于30%[28, 29, 30, 31]。最近, 已经发现了一些对草甘膦天然不敏感但具有接近于Ⅰ 类EPSPS的保守序列的新型EPSPS, 包括4G-1、GRG23、GRG51、AM79AroA、Gr5aroA和AroAJ.sp[32, 33, 34, 35, 36], 但是被批准商业化应用的草甘膦抗性基因却很少[18]。本研究使用的G10evo基因是浙江大学转基因抗虫植物和生物安全实验室独立克隆和改造获得的能够高抗草甘膦的新基因, 具有自主知识产权。其氨基酸序列与目前划分出的EPSPS家族Ⅰ 、Ⅱ 类抗草甘膦基因亲缘性很低, 保守序列相差很大, 既不属于Class Ⅰ 也不属于 Class Ⅱ [20, 21], 该基因在拟南芥(Arabidopsis thaliana)、烟草(Nicotiana tabacum)和大豆(Glycine max)中均表现出良好的草甘膦抗性, 用该基因培育的抗草甘膦大豆对草甘膦具有高抗性, 能够满足生产需要[22]。
我国转基因玉米新品种选育仍处于研发阶段。本研究以玉米泛素蛋白基因的启动子(pZmUbiquitin)作为G10evo基因的启动子, 同时以烟草花叶病毒 35S启动子 (35S promoter)作为增强子, 使用来源于玉米的乙酰羟基酸合酶(acetohydroxyacid synthase, AHAS)的信号肽, 利用农杆菌介导法将G10evo导入到玉米中, 筛选得到高抗草甘膦的转基因玉米AG16, 并从DNA、蛋白和功能性状三个方面对转基因玉米AG16进行分析检测。
DNA分子检测结果显示, G10evo基因单拷贝插入到AG16基因组中, T-DNA的单拷贝插入有利于外源基因的稳定遗传, 避免因为外源基因分离导致目的性状不稳定。Western blot结果显示, G10evo蛋白在AG16植株的根、茎和叶中均有表达, 外源蛋白大小与预期大小一致。ELISA定量分析结果显示, G10evo蛋白在AG16组织中均有较高水平的表达, 尤其是在嫩茎组织, 喷洒草甘膦前后, 表达量分别达到9.975和11.234 μ g· g-1。草甘膦抗性试验结果证明, AG16可以抗4~8倍田间用量的草甘膦(8.20~16.10 g· L-1)。抗除草剂是商业化转基因作物中最为重要的性状之一, 关于草甘膦抗性基因挖掘和应用的研究报道众多, 但是能够商业化应用的却很少。目前商业化应用的抗草甘膦主要有孟山都公司的CP4 EPSPS、先正达公司玉米EPSPS突变改良基因和GRG23, 其中又主要以CP4 EPSPS应用最为广泛。主要原因有以下几方面, 一是产权问题, 孟山都公司以CP4 EPSPS为中心, 申请了一系列专利保护, 将大多同源基因囊括在其保护群中; 二是抗性水平问题, 在生产应用中存在着重复喷施和农药错误配制的问题, 因此推广应用的抗草甘膦作物必须有较高的抗性, 能够降低用药错误造成药害的风险。本研究使用的抗草甘膦基因G10evo获得的抗草甘膦玉米转化系AG16中的外源T-DNA为单拷贝插入、遗传背景清晰, 抗草甘膦蛋白在玉米的各个组织中表达较高。AG16能够抗4~8倍田间施用量的草甘膦, 满足生产推广的需要, 为我国抗除草剂转基因玉米新品种培育提供了有价值的种质资源。
G10evo基因作为高抗草甘膦的基因, 除了应用于研发抗草甘膦大豆和玉米等作物中, 还可以用于培育抗草甘膦牧草, 如抗草甘膦苜蓿(Medicago sativa)。苜蓿被称为牧草之王, 其杂草的防治受时间、空间以及除草剂品种等多种因素的严格限制[37]。培育抗草甘膦苜蓿, 可以提高苜蓿杂草防治效率, 有效防止其它杂草对养分和空间的占用, 提高牧草青贮的质量和数量。抗草甘膦性状在草坪草上同样具有重要的应用价值, 在草坪养护过程中尤其是像高尔夫球场等高质量的草坪上, 对同种、属、科的其它杂草进行防除, 传统选择性除草剂的防治效果有限, 将抗草甘膦基因G10evo转入到草坪草如狗牙根(Cynodon dactylon)、结缕草(Zoysia japonica)、假俭草(Eremochloa ophiuroides)等常见草坪草中, 可以简化草坪杂草防治方法, 提高杂草防治效率。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
抗草甘膦转基因玉米AG16分子特征和抗性鉴定
刘苗苗
Molecular characterization and efficacy evaluation of a transgenic corn event AG16
Liu Miao-miao