{kind=link}
{kind=link}
飞行前后意大利蝗飞行肌及能源消耗比较
引用本文
窦洁, 张若燕, 刘敏, 钱雪, 肖宏伟, 季荣. 飞行前后意大利蝗飞行肌及能源消耗比较. 草业科学, 2017,34(8):1721-1726
Dou Jie, Zhang Ruo-yan, Liu Min, Qian Xue, Xiao Hong-wei, Roman Jashenko, Ji Rong. Comparison of flight muscle and energy consumption of pre- and post-flight in Calliptamus italicus(Orthoptera:Acrididae) . Pratacultural Science,2017,34(8): 1721-1726
Permissions
Dou Jie, Zhang Ruo-yan, Liu Min, Qian Xue, Xiao Hong-wei, Roman Jashenko, Ji Rong. Comparison of flight muscle and energy consumption of pre- and post-flight in Calliptamus italicus(Orthoptera:Acrididae) . Pratacultural Science,2017,34(8): 1721-1726
Copyright©2017, 《草业科学》编辑部
《草业科学》编辑部
飞行前后意大利蝗飞行肌及能源消耗比较
第一作者:窦洁(1992-),女,新疆奎屯人,在读硕士生,主要从事新疆草原虫害生物学研究。E-mail:doujie92@163.com
摘要
意大利蝗( Calliptamus italicus)具有长距离迁飞行为,并常给迁入区造成严重损失。若要提高对其监测预警和防控水平,就需要掌握其飞行肌结构特征,分析其体内能源物质积累与飞行消耗的动态,因此本研究比较了意大利蝗吊飞前后飞行肌肌节长度、肌原纤维直径和线粒体含量差异及飞行不同距离虫体内糖原和甘油酯含量变化。结果表明,吊飞组与未吊飞对照组雌虫之间、雄虫之间线粒体含量均差异显著( P<0.05)。意大利蝗雌成虫飞行超过2.8 km后糖原消耗最多,为11.15 mg·g-1,雄虫飞行2.12.8 km时消耗最多,为16.10 mg·g-1。雌、雄成虫均在飞行距离大于2.8 km后甘油酯消耗最多,分别为61.69和69.75 mg·g-1。意大利蝗飞行同时消耗糖类和脂肪两类能源物质,且雌雄成虫分别飞行超过2.8 km和2.12.8 km时糖原利用效率最大,飞行超过2.8 km后甘油酯利用效率最大。本研究得出吊飞组个体能改变自身的飞行肌结构来适应飞行活动;且飞行过程中意大利蝗能同时利用糖类与脂类,糖类能快速产生能量以供起飞与短距离飞行,随后主要消耗甘油酯供长距离飞行。
关键词:
意大利蝗; 线粒体含量; 飞行肌; 能源物质; 利用效率
中图分类号:Q969.26+5.1
文献标志码:A
文章编号:1001-0629(2017)08-1721-06
doi: 10.11829/j.issn.1001-0629.2017-0066
Comparison of flight muscle and energy consumption of pre- and post-flight in Calliptamus italicus(Orthoptera:Acrididae)
Abstract
Calliptamus italicus has the ability to fly long distance for migration, which can have negative effects in migrated areas. In this study on C. italicus, we compared the sarcomere length, myofibrils diameter, mitochondrial content, and changes in glycogen and glyceride levels during different flight distances. Our results indicate significant ( P<0.05) differences between females, males, females and males of test group members and control group in the mitochondrial content. The most glycogen consumption was recorded for females flying over 2.8 km and males flying between 2.12.8 km, which were 11.15 and 16.10 mg·g-1, respectively. The most glyceride consumed for both, females and males was to fly over 2.8 km, which were 61.69 and 69.75 mg·g-1, respectively. We found that C. italicus consumed both, carbohydrates and lipid at the same time during flight. Females and males had the highest glycogen utilization efficiency when the travelling distances were over 2.8 and 2.12.8 km, respectively. Highest glyceride utilization efficiency was also observed when the travelling distance was over 2.8 km.
Keyword:
Calliptamus italicus; mitochondria content; flight muscle; energy substances; utilization efficiency
昆虫迁飞行为既是种群在进化过程中长期适应环境的遗传特性, 也是昆虫对栖息地资源发生季节性或偶然性变化时进化出的一种生存对策[1], 迁飞有助于昆虫寻找适宜的生境, 增加种群生存繁衍的机会[2, 3]。
飞行肌对昆虫飞行有重要意义, 肌节短、肌原纤维粗且飞行肌内线粒体含量多有利于肌肉快速收缩和远距离迁飞[4]。昆虫飞行消耗的能源物质种类包括脂肪、碳水化合物、氨基酸或者其中的两种或多种[5, 6, 7], 多数昆虫飞行时利用碳水化合物和脂肪供应能量。双翅目和膜翅目中的大多数昆虫远距离飞行以糖原作为能源物质[8, 9]; 直翅目和鳞翅目昆虫远距离飞行则主要消耗脂肪, 同翅目昆虫则以碳水化合物和脂肪两类物质同时作为能源供给[10]。东亚飞蝗(Locusta migratoria manilensis)在迁飞过程中将糖类和脂质同时作为能源物质, 脂肪体中的甘油激酶将甘油转变为海藻糖后直接进入糖酵解循环, 为飞行提供能量[6-10]。非洲粘虫(Mythimna separata)在飞行过程中, 糖类为成虫羽化及短时飞行提供能源[11], 脂类则在远距离迁飞中起重要作用。掌握昆虫迁飞能源物质消耗与利用动态有助于阐明昆虫迁飞机制, 为迁飞性害虫防治提供科学依据和决策参考。
意大利蝗(Calliptamus italicus)具有长距离迁飞行为, 属直翅目(Orthoptera)斑腿蝗科(Acridoidea)星翅蝗属(Calliptamus)。中国和哈萨克斯坦边境相邻的荒漠半荒漠草原是意大利蝗严重发生区, 借助有利的气象条件, 意大利蝗常跨境迁飞至新疆边境区域危害, 并造成了严重损失[12, 13]。由于意大利蝗迁飞危害具有突发性、暴发性和毁灭性, 为提高对其监测预警和防控水平, 掌握意大利蝗飞行肌结构特征, 本研究分析其迁飞能源物质消耗, 以期为预测中哈边境相邻区域意大利蝗种群迁飞动态及防控提供科学参考。
1 材料与方法
1.1 试虫饲养与吊飞处理
2016年7月于新疆维吾尔自治区昌吉玛纳斯南山蝗区(43° 54'N、86° 07'E, 海拔1 310 m)采集意大利蝗末龄蝗蝻, 将雌雄蝗蝻分开饲养于虫笼中(95 cm× 95 cm× 95 cm), 虫口密度70100头· m-2以获得群居型意大利蝗成虫。群居型意大利蝗判断采用张洋[14]的方法, 首先根据其体色及前胸背板, 群居型意大利蝗体色比散居型深, 且前翅有两条明显白色长条纹; 其次依据前翅长度与后足股节宽度比值, 如介于1.421.94为群居型, 小于1.42则为散居型[14]; 每天以新鲜玉米(Zea mays)、小麦(Triticum aestivum)、苜蓿(Medicago sativa)和冷蒿(Artemisia frigida)混合饲喂, 并以麦麸辅助饲养。在(25± 3) ℃、湿度为(50%± 5%)时, 自成虫开始羽化逐日选取健康且体重相近(± 0.2 g)的同性别成虫吊飞(JDFXM-3, 佳多), 每次吊飞24头(♀∶ ♂=1∶ 1), 重复3次, 以获得雌雄成虫飞行能力最强的日龄。根据飞行距离得出7日龄雌虫和2日龄雄虫的迁飞能力最强。
1.2 飞行前后飞行肌结构测量
将飞行能力最强日龄的雌、雄成虫吊飞10 h(10:00-20:00)后选取健康个体在0.65%生理盐水中解剖其飞行肌, 戊二酸固定, 环氧树脂包埋聚合(EPON872包埋), 超薄切片(Leica uc6)70 nm, 电镜下(JEOL 1230, 日本电子株式会社)观察并测量相邻两个Z带之间的距离即肌节长度(nm)、肌纤维直径(nm)和线粒体在肌纤维所占的体积百分含量(%)。每次吊飞24头(♀∶ ♂=1∶ 1), 重复3次, 以未吊飞的相同日龄和性别且体重相近(± 0.2 g)的成虫作为对照组。
肌节长度和肌原纤维直径在电子显微镜下测量, 线粒体含量用其所占方格纸格数除以总格数得到。
1.3 飞行前后糖原和甘油酯测定
将飞行能力最强日龄的雌、雄成虫吊飞10 h(10:00-20:00)后按照不同飞行距离进行分组, 并于液氮罐中处死置于-80 ℃冰箱保存。剔除吊飞过程中死亡或飞行距离≤ 100 m的个体, 以未吊飞的相同日龄和性别且体重相近(± 0.2 g)的成虫作为对照组。
糖原含量采用蒽酮比色法[15]测定, 甘油酯含量采用甘油酯酶法测定, 甘油酯试剂盒(Tissue triglyceride assay kit E1013)由普利莱基因技术有限公司(北京)提供。
能源消耗量(mg· g-1)指对照组体内能源物质含量减去吊飞组体内能源物质含量。
利用率[mg· (km· ind)-1]是指虫体内能源物质含量与飞行距离和试虫头数乘积的比值, 可用来评价昆虫飞行能源利用效率情况, 利用率越小表明个体单位飞行距离所需的能源物质越少, 说明能源利用效率越高。
1.4 数据处理
意大利蝗最强飞行日龄雌雄成虫之间肌节长度、肌原纤维直径、线粒体含量差异采用t-检验(P< 0.05); 吊飞前后意大利蝗体内糖原、甘油酯含量采用单因素方差分析(One-way ANOVA), 结合最小显著差数法(LSD)和多范围检验(Duncan’ s检验)分析差异显著性(P< 0.05), 数据分析软件采用SPSS 19.0。
2 结果与分析
2.1 吊飞组和对照组飞行肌结构比较
吊飞组雌、雄成虫的肌节长度分别为2 939.9和2 487.47 nm, 对照组分别为3 171.43和3 545.47 nm; 吊飞组雌、雄成虫的肌原纤维直径分别为1 134.28和1 268.03 nm, 对照组分别为905.04和749.61 nm; 两组雌虫之间、雄虫两组之间的肌节长度、肌原纤维直径无显著差异(P> 0.05); 吊飞组雌、雄成虫的线粒体含量分别为36.07%和35.86%, 对照组分别为23.91%和22.38%, 两组雌虫之间、雄虫之间差异显著(P< 0.05)(表1)。
| 表1 吊飞前后意大利蝗雌雄成虫肌节长度、肌原纤维直径、线粒体含量比较 Table 1 Sarcomere length, myofibrils diameter and mitochondria content of females and males Calliptamus italicus adults of pre- and post-flight |
2.2 飞行前后能源物质消耗比较
根据吊飞组意大利蝗雌雄成虫的飞行距离分为5组, 即< 0.7、0.71.4、1.42.1、2.12.8和≥ 2.8 km。随飞行距离增加, 雌虫体内糖原消耗量增加, 飞行距离超过2.8 km后体内糖原消耗最多, 为11.15 mg· g-1, 显著高于飞行距离< 0.7、0.71.4 km的糖原消耗(P< 0.05); 雌虫飞行距离< 0.7和0.71.4 km之间、1.42.1和2.12.8 km之间差异不显著(P> 0.05)。雄虫体内糖原消耗量则随飞行距离增加先增加后减少, 飞行距离在2.12.8 km时糖原消耗最多, 为16.10 mg· g-1, 显著高于飞行距离< 0.7、0.71.4和≥ 2.8 km的糖原消耗量; 飞行距离1.42.1 km的显著高于< 0.7和0.71.4 km的糖原消耗量; 雄虫飞行距离< 0.7、0.71.4 和≥ 2.8 km之间糖原消耗差异不显著(P> 0.05)(图1)。飞行距离1.42.1、2.12.8和≥ 2.8 km时雌、雄成虫之间糖原消耗差异显著(P< 0.05)。
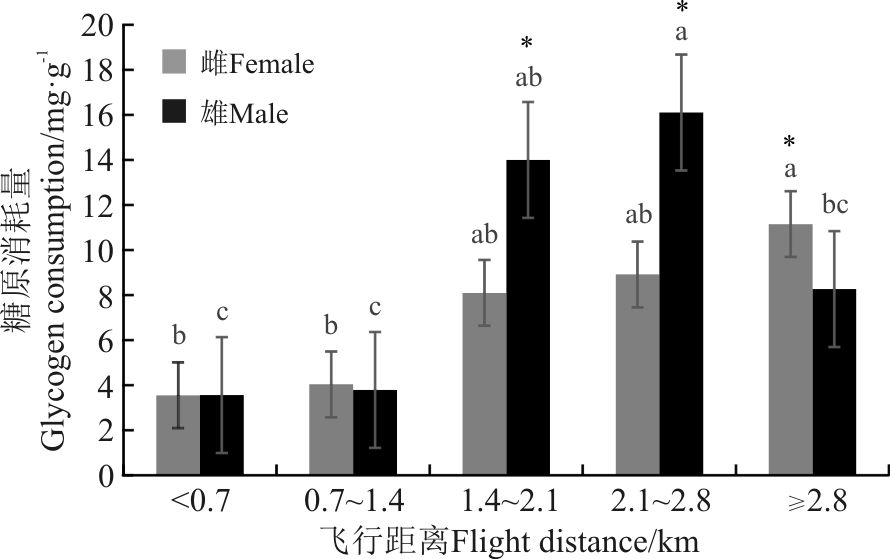 | 图1 不同飞行距离意大利蝗糖原消耗比较 注:不同小写字母表示同一性别(雌虫、雄虫)不同飞行距离间差异显著(最小显著差数法LSD和Duncan’ s检验, P< 0.05); * 表示同一飞行距离雌虫和雄虫间差异显著( P< 0.05) 。图2同。Fig. 1 Glycogen consumption of different flight distance of Calliptamus italicus Note:Different lowercase letters indicate significant difference of males or femals between different flight distances at the 0.05 level; * for the same flight distance indicate significant difference between females and males at the 0.05 level; similarly for Fig. 2. |
随飞行距离增加, 雌雄成虫甘油酯消耗量逐渐增加, 飞行距离超过2.8 km后雌雄成虫体内甘油酯消耗量最多, 分别为61.69和69.75 mg· g-1(图2)。雌虫两组之间、雄虫两组之间均在飞行距离≥ 2.8 km时显著高于< 0.7 km时的甘油酯消耗量(P< 0.05); 飞行距离为0.71.4、1.42.1和2.12.8 km时甘油酯消耗量与< 0.7和≥ 2.8 km时的甘油酯消耗量均无显著差异(P> 0.05)。任何飞行距离条件下, 雌雄成虫之间无显著差异(P> 0.05)。
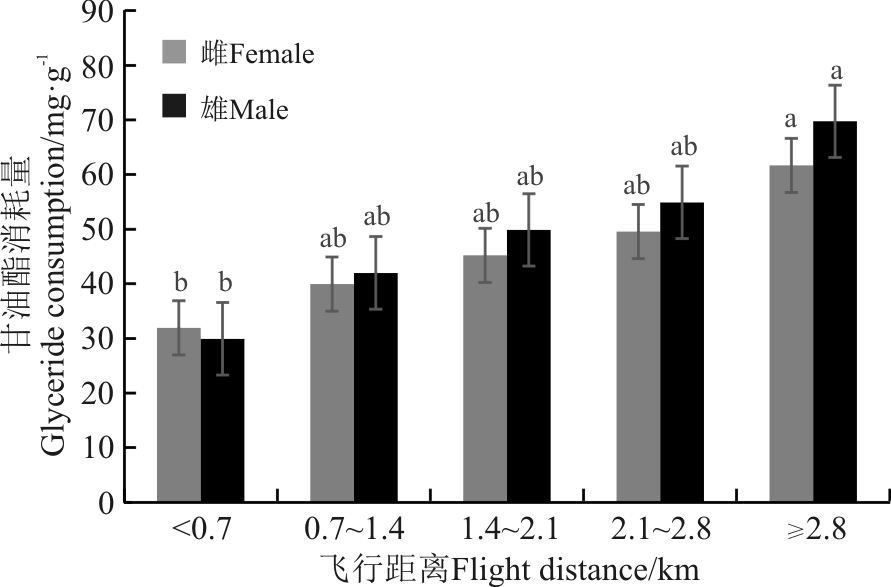 | 图2 不同飞行距离意大利蝗甘油酯消耗比较Fig. 2 Glyceride consumption of different flight distance of Calliptamus italicus |
2.3 意大利蝗飞行中体内糖原和甘油酯利用率分析
意大利蝗雌雄成虫飞行距离分别在≥ 2.8和2.12.8 km时, 糖原利用率最小, 表明单位飞行距离糖原消耗最少, 利用效率最高; 飞行距离≥ 2.8 km, 雌、雄成虫甘油酯利用率均最小, 表明单位飞行距离甘油酯消耗最少, 利用效率最高。飞行距离对雌虫、雄虫的糖原和甘油酯的利用率均有显著影响(P< 0.05), 相同飞行距离下, 成虫的性别对糖原和甘油酯的利用率均无显著影响(P> 0.05)(表2)。
| 表2 意大利蝗飞行后糖原和甘油酯利用率比较 Table 2 Glycogen and glyceride utilization efficiency of Calliptamus italicus after flight |
3 讨论与结论
昆虫的飞行能力很大程度上取决于其飞行肌结构的发育情况[16]。肌原纤维和线粒体是飞行肌中的重要组成部分, 也是昆虫飞行生理系统的动力来源中心[17], 线粒体为肌原纤维收缩提供能量, 肌原纤维为昆虫飞行提供动力[18]。在研究烟草天蛾(Manduca sexta)、天蚕蛾(Antheraea yamamai)、粘虫(Mythimna separate)和东亚飞蝗时发现, 肌节短、肌原纤维直径粗且线粒体含量多有利于昆虫长距离飞行, 因较短的肌节长度和粗的肌纤维直径能够使肌肉收缩频率加快, 使其能高频振翅, 同时更多的线粒体含量保证提供更多的能量以供飞行[19, 20]。本研究亦得出相似结论, 吊飞组个体的飞行肌结构更适于飞行。为了比较飞行前后飞行肌和能量消耗差异, 本研究只选择了飞行能力最强日龄雌雄成虫作为研究对象, 但逐日龄成虫飞行肌及飞行能量消耗仍是值得探讨的问题。
昆虫飞行主要利用的能源有糖类、脂类和氨基酸。糖类能快速产生能量, 因而是多数昆虫起飞或短距离飞行主要能源; 脂类因能量较高, 适合昆虫长距离的飞行。本研究得出, 意大利蝗飞行消耗的能源物质种类包括糖类和脂类, 这与东亚飞蝗、亚洲小车蝗(Oedaleus asiaticus)[21]的研究结果一致。研究发现, 随飞行距离增加意大利蝗、雄虫的糖原利用效率先增后略微下降, 雌虫的糖原利用效率则逐渐增加; 随飞行距离增加雌、雄成虫甘油酯利用效率均增大, 且飞行不同距离糖原利用效率均大于甘油酯。究其原因, 这与飞行起始过程阶段主要消耗糖类, 能快速产生能量以供起飞与短距离飞行, 随后主要消耗甘油酯供长距离飞行有关。飞行不同距离意大利蝗雄虫体内糖原和甘油酯消耗均大于雌虫, 这可能与雌雄生殖活动有关。意大利蝗成虫在羽化16 d左右开始交配[22], 雄虫最强飞行日龄为2日龄, 推测此时更多能量消耗用于飞行寻找合适交配繁殖的雌虫, 而雌虫最强飞行日龄为7日龄, 此时更多能量消耗用于卵巢发育。研究发现, 同多数迁飞性昆虫一样, 群居型意大利蝗较散居型有更强的飞行能力, 前者最大飞行距离可达2.66 km, 后者仅为0.2 km[23], 但群居型意大利蝗借助气流其扩散距离可达120 km[24]。因此, 中哈边境蝗虫跨境迁飞需借助当地盛行的西北风才能实现远距离迁飞[25]。本研究结果可为在中国和哈萨克斯坦边境区域蝗虫迁飞路径中建立监测预警平台提供决策依据。
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|