
{kind=link}
{kind=link}
吐鲁番沙虎消化道对刺山柑种子吸水和萌发的影响
[林英英 , 肖丽蓉, 马晓燕, 时磊]
, 肖丽蓉, 马晓燕, 时磊]
, 肖丽蓉, 马晓燕, 时磊]
|
|


第一作者:林英英(1987-),女,河南宝丰人,在读硕士生,主要从事动物生态学研究。E-mail:ly1987@126.com
取过吐鲁番沙虎( Teratoscincus roborowskii)消化道和直接从果实分离的刺山柑( Capparis spinosa)种子为供试材料,4 ℃预冷处理60 d后转入25 ℃恒温培养箱进行为期30 d的萌发试验,研究过吐鲁番沙虎消化道对刺山柑种子吸水能力和萌发的影响。结果表明,过吐鲁番沙虎消化道对种子的前期吸水能力和萌发有显著影响( P<0.05),消化作用增强了种子的快速吸水能力,同时提高了萌发率,但对种子的萌发速度和生活力无显著影响( P>0.05)。
In this study, seeds of caper ( Capparis spinosa) separated from feces of Turpan wonder gecko ( Teratoscincus roborowskii) were used as the treatment groups, and seeds separated from fresh fruit were used as the control groups. The treatment and control groups were both precooled at 4 ℃ for 60 days and then transferred to germination experiment in a constant temperature of 25 ℃ incubator for 30 days. The effects of passage through Turpan wonder gecko digestive tracts on caper seed’s water uptake ability and germination were determined. The results showed that seed through the digestive tract of Turpan wonder gecko had significant impacts ( P<0.05) on water uptake ability and germination, the digestive function promoted the seed’s fast water uptake ability and improved seed germination percentage, but did not affect ( P>0.05) germination rate and viability of caper seed.
动物的种子扩散作用是动植物相互关系研究的一个重要方向, 其扩散作用主要表现在肉质果类植物中[1], 鸟类和哺乳类是参与这类扩散最常见的动物类群[2]。过食果动物消化道对种子萌发能力的影响是衡量扩散质量的重要指标。食果动物促进种子萌发的一个主要机制是消化道对种皮的研磨作用[2], 这种作用导致种子对水和气体的渗透性增强[3]。鸟类和哺乳类对肉质果植物的种子扩散是广为人知的, 但关于爬行类动物的种子扩散研究数据还较少, 现有的研究发现, 过爬行类动物消化道对41种(56%)植物种子的萌发没有影响, 且促进(28%)作用多于抑制作用(16%)[4]。一般认为, 蜥蜴(Lizard)的种子扩散作用是不重要的[5], 可能是因为大部分蜥蜴是肉食性的[6]。然而, 最近的研究表明, 植食性蜥蜴可以影响植物的繁殖和群落结构[7]; 有些蜥蜴还和植物建立了紧密的联系, 对花和果实的进化也产生了选择压力[8]。
一些研究发现, 经过蜥蜴的消化道后种子的萌发得以促进[7]。植物的种皮和果皮等结构, 虽然对胚有保护和营养等功能, 但它们的存在使胚得不到充足的水分和氧气, 对萌发过程有抑制作用。蜥蜴的消化道对种皮的碾磨增加了其对水和空气的渗透性从而促进萌发[9]。与直接从果实分离的种子相比, 过普通壁虎(Hoplodactylus maculates)消化道后种子萌发率增加, 表明普通壁虎是有效的种子扩散者[10]。通过栉捷蜥(Liolaemus bellii)的消化道后, 安第斯灌丛植物岩高兰小檗(Berberis empetrifolia)种子的萌发率大于未过消化道的对照组[11]。
刺山柑(Capparis spinosa)为散生灌木, 根深粗壮, 主茎不明显, 匍匐或悬垂, 花大, 白色, 单生于叶腋, 在4月下旬或5月上旬开始有花, 花期可长达4个月。从开花到果熟约需30 d, 蒴果浆果状, 椭圆形至倒卵形, 成熟后淡红色, 干后常四裂, 种子肾形, 直径约3 mm, 多数褐色[12]。自然状态下, 刺山柑种子萌发率很低。成熟后2~3个月刺山柑种子的萌发率只有5%[13]。推测萌发率较低是由种皮外覆盖物引起休眠造成的, 因为使用浓H2SO4处理种子有利于刺山柑种子萌发[14]。刺山柑极耐旱、耐高温和抗风蚀, 具有保持水土、防风固沙和保护绿洲及工程设施的生态功能, 是恶劣、严酷生境造就出来的一种高经济价值、生态功能显著的荒漠植物[15]。
吐鲁番沙虎(Teratoscincus roborowskii Bedriaga, 1905/1906), 分布于中国新疆的吐鲁番地区, 为中国特有种[16]。其在夏季采食大量的刺山柑果实, 从其粪便中可分离出大量完整的刺山柑种子, 这种现象在北半球温带荒漠地区的壁虎中首次报道, 同时在整个广义壁虎类(Gekkota)动物中也是十分罕见的[17]。据此推测吐鲁番沙虎可能是刺山柑种子的扩散者。由此提出如下假设:1)过吐鲁番沙虎消化道能提高刺山柑种子的吸水能力; 2)过吐鲁番沙虎消化道促进刺山柑种子的萌发进程。本研究旨在定性分析过消化道对种子吸水和萌发的影响, 探究吐鲁番沙虎是否是刺山柑种子的有效扩散者。
研究地位于中国科学院吐鲁番沙漠植物园, 地处吐鲁番盆地东南部恰特喀勒乡境内, 距吐鲁番市东南约10 km, 距乌鲁木齐200 km, 89° 11' E, 40 ° 51' N, 海拔-95~-76 m[18]。野外工作时间为2014年8月。
2014年8月, 在吐鲁番沙漠植物园捕捉吐鲁番沙虎82只, 禁食并单独暂养在养殖盒中, 及时收集含刺山柑种子的粪便, 用蒸馏水分离该粪便中的刺山柑种子。采集新鲜成熟的刺山柑果实, 手工去除果肉, 蒸馏水洗涤种子。分离出的过消化道和未过消化道(对照组)种子风干后放在牛皮纸信封中, 常温干燥环境下保存1个月。
取过消化道种子和未过消化道的种子各480粒, 160粒为1个重复, 3组重复。放入4 ℃恒温箱里, 预冷处理60 d, 以打破休眠[19]。
取过吐鲁番沙虎消化道和对照组的刺山柑种子各120粒, 3组重复, 每组重复40粒。吸水试验前先用电子天平(1/10 000 g)称取种子初始重, 然后将各处理种子分别置于培养皿中, 加入常温蒸馏水分别浸泡2、4、6、8、10和12 h, 在滤纸上吸干种子表面水分, 称质量并记录6个吸水时间段的种子重。由此计算每处理组种子6个吸水时间段的种子吸水率(Rn)[20]。
Rn=(mn-m0)/m0× 100% .
式中:m0为浸种前种子的质量; mn为浸种n时间后种子的质量, n为浸种时间(h), n=2, 4, 6, 8, 10, 12。
1.5.1 萌发试验材料及方法 萌发试验所需要的培养皿和镊子均用高压蒸汽消毒。然后把种子按组分别摆放在铺有两层滤纸的培养皿中, 注入0.2% KNO3溶液, 达到饱和为止。置于25 ℃恒温培养箱, 12 h光照/12 h黑暗条件下进行种子萌发试验。萌发以种子露白为准, 每天检查一次萌发情况, 滴加0.2% KNO3溶液, 保持培养皿内滤纸的湿润。30 d后结束萌发试验, 统计数据, 计算萌发率, 此处衡量萌发速度的指标采用平均萌发天数[19]、开始和最后萌发天数。
1.5.2 萌发数据计算公式
萌发率=(萌发种子总数/有生活力种子总数)× 100%;
平均萌发天数=(n1× t1+n2× t2+n3× t3+…+ni× ti)/T。
式中:ni为萌发种子的天数, ti为每天的萌发种子数, T为萌发的种子总数。
用SPSS 18.0软件进行数据分析。统计检验前对数据进行方差齐性检验, 萌发率及种子生活力数据方差不齐(P< 0.05), 对其进行反正弦平方根转换使其符合正态分布, 然后采用独立样本T检验对刺山柑种子吸水率、萌发率、种子生活力、平均萌发天数、开始萌发的天数和最后萌发的天数数据进行检验, 显著性水平设置为α =0.05。
浸种2-6 h时, 过吐鲁番沙虎消化道的种子和对照组种子的吸水率差异显著(P< 0.05), 浸种8-12 h时二者差异不显著(P> 0.05)(图1)。过消化道和对照组的种子, 在浸种0-2 h达到较高的吸水率, 吸水速度最高; 4-6 h吸水率升高缓慢, 吸水速度降低; 浸种8-12 h吸水率基本恒定, 吸水速度最低。不同的浸种时间, 过消化道组种子的吸水率始终高于对照组, 说明过吐鲁番沙虎消化道对种子的吸水起促进作用。
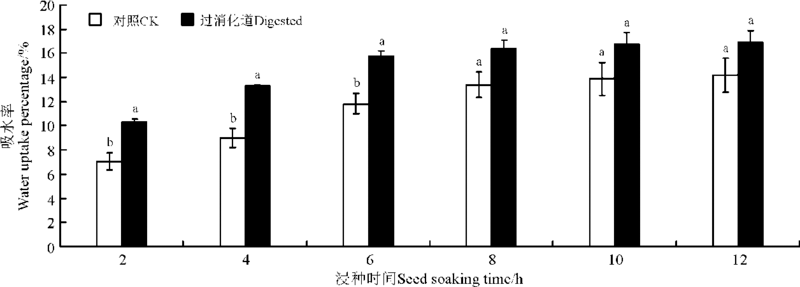 | 图1 过吐鲁番沙虎消化道和对照组刺山柑种子在不同浸种时间的吸水率 注:不同小写字母表示相同浸种时间不同处理间差异显著(P< 0.05)。Fig.1 Water uptake percentage of caper seeds which had passed through the digestive tract of Turpan wonder gecko and control groups in various seed soaking time Note: Different lower case letters show significant difference at same seed soaking time between the two treatments at 0.05 level. |
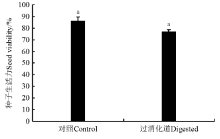
过消化道和对照组刺山柑种子的生活力差异不显著(P=0.115> 0.05)(图2)。说明过吐鲁番沙虎的消化道对刺山柑种子的生活力没有影响。
 | 图2 过吐鲁番沙虎消化道组和对照组刺山柑种子的生活力 注:相同小写字母表示不同处理间差异不显著(P> 0.05)。Fig.2 Seed viability of caper seeds that had passed through the digestive tract of Turpan wonder gecko and control groups Note: Same lower case letters show no significant difference between the two treatments at 0.05 level. |
过消化道组种子的30 d萌发率(9.73%)显著高于对照组的(2.92%)(P=0.035< 0.05)。但, 过消化道和对照组种子的平均萌发天数(P=0.590> 0.05)、开始萌发天数(P=0.423> 0.05)以及最后萌发天数(P=0.386> 0.05)差异不显著。
种子萌发成功与否与其萌发前的吸水过程密切相关, 大部分植物种子都具有相同的吸水规律, 包含快速吸水、缓慢吸水和稳定吸水3个阶段[22]。刺山柑种子也有相似的吸水规律:0-2 h快速吸水, 达到较高的吸水率, 4-6 h吸水率增加缓慢, 8-12 h呈稳定趋势, 所以0-2 h、4-6 h、8-12 h分别是刺山柑种子的快速吸水阶段、缓慢吸水阶段和稳定吸水阶段。禾本科植物的种子在4-5 h之前为种子的快速吸水阶段, 藜科植物种子在3 h之前为种子的快速吸水阶段, 豆科植物种子在5-6 h之前为种子的快速吸水阶段[22]。相比之下, 刺山柑种子的快速吸水阶段时间比较短(0-2 h)。本研究中在刺山柑种子的快速吸水和缓慢吸水两个阶段, 过吐鲁番沙虎消化道种子的吸水率均显著高于对照组, 说明过沙虎消化道提高了刺山柑种子在这两个阶段的吸水速度。在种子吸水的整个过程中, 过消化道种子的吸水率也始终高于对照组, 说明过沙虎消化道促进了种子吸水。在干旱、半干旱区, 水分是影响种子萌发及生长的一个限制因子[23]。吐鲁番地区常年干旱少雨, 吐鲁番沙虎消化道对刺山柑种子吸水能力有促进作用, 这对荒漠植物刺山柑在当地的植被更新有重要意义。
| 表1 刺山柑种子的萌发率及萌发速度 Table 1 Germination percentage and germination rate of caper seeds |
消化意味着对种皮的化学处理或机械研磨, 这可能抑制或促进种子萌发。有学者发现, 蜥蜴的消化过程不促进种子萌发, 甚至对某些植物种子的萌发有负面作用[9, 24]。然而另外一些物种, 特别是产核果或浆果类的肉质果, 经过蜥蜴消化道后萌发率通常会提高[7, 25]。本研究表明, 经过吐鲁番沙虎消化道后刺山柑种子的萌发率显著高于对照组。但是过消化道和对照组的萌发率都不高, 这可能是由刺山柑种子较厚的种皮对种胚获得氧气和水的限制作用导致的。GA3可以提高刺山柑种子的萌发率, 最高可达60%[14]。本研究未采用GA3处理种子, 所得到的萌发结果更接近野外的实际情况。除了消化道对种子萌发的促进作用, 吐鲁番沙虎夏季食物组成中超过80%是刺山柑果实[17], 由此可以扩散大量的刺山柑种子离开母株[26], 并到达适合种子萌发和幼苗建植的微生境[27]。蜥蜴的消化过程可以提高种子生活力[7], 或降低种子生活力[7], 但大多数植物种子生活力不受影响[27, 28], 本研究发现过消化道对刺山柑种子的生活力没有影响。
在恶劣环境条件下生长的植物会在长期进化过程中形成两种对立的萌发对策, 一是种子成熟后在环境适宜的条件下仍保持休眠而保留在种子库中, 从而降低了种子萌发及幼苗定植过程中的死亡风险; 二是种子一旦遇到适宜的环境条件(比如光照、水分或温度等)就会立即萌发, 种子快速萌发而占领生境[29]。河西走廊荒漠地带的枸杞(Lycium barbarum)、黑果枸杞(L. ruthenicum)、花棒(Hedysarum scorpaium)和泡果白刺(Nitraria sphaerocarpa)4种旱生灌木种子很可能采取第二种萌发对策, 它们可在适宜的条件下短时间内达到萌发高峰[30], 这种萌发对策是长期生活在干旱、高温地带的荒漠植物适应环境的结果[31]。萌发速度是影响幼苗成活和植被重建的关键因素, 在种子扩散阶段, 动物的消化作用可能影响种子的萌发速度。有研究发现, 爬行动物对47%的植物种子萌发速度起促进作用, 37%没有影响, 剩余16%有抑制作用[4]。本研究发现, 过吐鲁番沙虎消化道的刺山柑种子的平均萌发天数、开始萌发天数及最后萌发天数均与对照差异不显著, 表明沙虎消化道对刺山柑种子的萌发速度没有影响。刺山柑种子萌发率低且萌发速度较慢, 表明其存在休眠现象, 有利于形成土壤种子库。干旱或半干旱地区的植物可能通过保持长时间连续萌发, 从而降低环境波动带来的风险[32]。
本研究结果支持过吐鲁番沙虎消化道提高刺山柑种子的吸水和萌发能力的假设, 即定性地证明了吐鲁番沙虎是刺山柑种子的潜在扩散者。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|