{kind=link}
德阳市3种林草模式下土壤动物群落特征
[吴韬 , 冯茂松, 韩东苗, 李文兵, 铁烈华]
, 冯茂松, 韩东苗, 李文兵, 铁烈华]
, 冯茂松, 韩东苗, 李文兵, 铁烈华]
|
|


第一作者:吴韬(1992-),男,重庆云阳人,在读硕士生,主要研究方向为人工林造林理论及技术。E-mail:329801466@qq.com
为了解林草模式下土壤动物组成及群落结构特征,以核桃( Juglans regia) +紫花苜蓿( Medicago sativa)、核桃 +菊苣( Cichorium intybus)及核桃 +鸭茅( Dactylis glomerata)3种模式为研究对象,同时以纯核桃林模式作为对照进行土壤动物调查。结果表明,1)调查共获土壤动物303只,分属3门7纲17目,优势类群为线虫纲(Nematoda)、膜翅目(Hymenoptera)、蜱螨目(Acarina);2)林草模式下土壤动物表聚性明显,纯林模式下土壤动物个体密度土层分布表现为5-10 cm>10-15 cm>0-5 cm;3)核桃+苜蓿模式下Shannon-Wiener多样性指数和密度-类群指数(DG)最高,纯林模式下密度-类群指数(DG)最低,但Pielou均匀度指数最高;4)核桃+苜蓿模式下虽然土壤动物个体数明显高于纯林模式,但在类群种类上却与纯林模式最为相似;5) 4种模式中,核桃+苜蓿模式腐食性(Saprozoic)土壤动物种团比例最高。综合而言,3种林草模式均能有效增长土壤动物群落组成,尤其以核桃+苜蓿模式土壤动物群落丰富程度最高。
This study was conducted to study the composition and community structure of soil fauna in three types of forest-grass patterns ( Juglans regia+Medicago sativa, Cichorium intybus and Dactylis glomerata), and a pure Juglans woodland was applied as a control group in Deyang, Sichuan Province. The results showed that, A total of 303 specimens of soil faunas was found in this study, which were belonged to 3 phylum, 7 classes and 17 orders. Of those, Nematoda, Hymenoptera and Acarina were the dominant groups. The soil fauna was found to gather on the surface of earth in three types of forest-grass pattern, while the order of soil fauna density was 5-10 cm>10-15 cm>0-5 cm. In addition, the diversity of Shannon-Wiener index and DG (Density-groups) index of soil fauna was the highest in the J. regia+M. sativa pattern while the lowest DG index in pure woodland that possessed the highest Pielou index; The J. regia+M. sativa pattern has a significant difference to the pure woodland pattern in individual number of soil fauna, but on the area of species of groups are similar to the pure woodland pattern, and it had a highest proportion of Saprozoic fauna in four patterns. In summary, the three forest-grass patterns can enrich the composition of soil fauna communities, and highest abundant soil fauna was observed in J. regia+M. sativa pattern.
土壤动物作为生态系统中重要的组成部分, 不仅能够改善土壤结构, 参与物质、能量的循环, 同时由于其数量庞大、种类繁多、活动范围宽广、对土壤环境变化敏感等特点, 常被作为反映土地健康状况的指示性生物[1, 2, 3, 4]。林草模式是农林复合系统(Agroforestry System)的一个类型, 将多年生木本植物与农业、牧业在同一土地单位不同空间有机结合的复合型经营模式[5], 通过高效的空间结构形成物质与能量的可持续循环, 提高生产力, 充分发挥林地生态效益和经济效益。核桃(Juglans regia)树种经济效益显著, 大力发展核桃产业可转移农村剩余劳动力, 增加农民收入, 带动农产品及相关产业的发展。
目前, 关于森林土壤动物的研究多集中于典型森林生态系统内的群落特征[6, 7, 8], 不同土地利用方式下的群落关系[9, 10, 11]等方面, 有关土壤动物在农林复合生态系统中群落多样性研究鲜有报道。为此, 于2014年9月对核桃分别复合紫花苜蓿(Medicago sativa)、菊苣(Cichorium intybus)及鸭茅(Dactylis glomerata)而成的3种林草模式下土壤动物群落进行初步调查分析, 并以纯核桃林模式为对照, 旨在观察了解土壤动物群落在复合系统中的表现, 总结分析林草模式与纯林模式下土壤动物多样性特点, 为该地区选择最适林草复合模式, 进而改善农村产业结构、促进农村经济发展提供新思路和新技术。
试验地位于四川省德阳市旌阳区和新镇永新村(104° 25'30''-104° 25'45'' E, 31° 04'09''-31° 04'15'' N), 属湿润季风气候, 年平均气温16.4 ℃, 最高气温36.5 ℃, 最低气温-6.7 ℃, 年平均降水量900 mm, 年平均风速1.5 m· s-1, 无霜期长达276 d。土壤主要为紫色土, 土层较浅, 呈中性或微碱性反应, 有机质含量低。地带性植被为柏木(Cupressus funebris)人工林和天然次生林。
于2011年对试验地内柏木进行带状或块状皆伐, 在前期整地时采用局部深耕整地方式, 并施用以有机肥为主的基肥, 整地完成后, 次年以3 m× 4 m的品字形排列种植一年生嫁接核桃苗, 并做好松土除草、施肥、灌溉与排水等幼林地抚育工作, 同时设立3个随机区组, 区组内环境条件相对一致, 每个区组划分为4个小区, 包含核桃+苜蓿(模式Ⅰ )、核桃+菊苣(模式Ⅱ )、核桃+鸭茅(模式Ⅲ )3种林草模式以及作为对照的核桃纯林模式(模式Ⅳ ), 共12个样地。2014年9月在上述每个样地内, 用典型选样的方法随机布设3个取样点, 除去地表杂草苔藓等覆盖物, 挖取50 cm× 50 cm面积土样, 分0-5、5-10和10-15 cm土层进行手捡计数, 收集大型土壤动物并放入盛有75%酒精的容器内带回室内鉴定; 同时, 根据相同方法随机布设3个取样点收集中小型土壤动物, 分别用100和25 mL的圆形取样器按0-5、5-10和10-15 cm连续分层采取干生土壤动物和活动性较弱的湿生土壤动物。取好的土样须迅速用0.15 mm的尼龙纱包裹好放入写有标签的黑色小布袋中, 以防止土壤动物逃逸, 带回室内分离鉴定。土壤动物的分离均在烘虫箱中进行。在中小型干生土壤动物的烘虫箱中放置直径为10 cm的漏斗, 筛孔直径为1 mm; 在中小型湿生土壤动物的烘虫箱中放置直径为10 cm的漏斗, 下接夹有两个止水夹的乳胶管, 筛孔直径为0.25 mm。两种烘虫箱的温度控制在35~40 ℃, 烘虫时间均为48 h, 分离出的干生土壤动物用盛装75%酒精的培养皿收集, 在解剖镜下观测土壤动物数量和种类, 收集湿生土壤动物的容器不加酒精, 直接在清水中用解剖镜观察鉴定种类和数量, 干生动物每12 h观测一次, 湿生动物开始每4 h观测一次, 以后时间间隔逐步加长, 以防止线蚓自溶[12]。
类群数量等级划分:个体数量占个体总数10%以上的为优势类群, 占个体总数1%~10%的为常见类群, 少于1%的为稀有类群。
多样性指数:选用Shannon-Wiener多样性指数(H)、Simpson优势度指数(C)、Pielou均匀度指数(J)和密度-类群指数(Density-Groups, DG)来描述土壤动物的群落状况, 公式如下:
Shannon Wiener指数:H=-
Simpson指数:C=
Pielou指数:J=H/lnS;
Density-Groups:DG=(g/G)
式中, S为样方中类群个数; Pi=ni/N; ni为第i个类群的个体数目, N为所有类群的个体数目, Di为第i个类群的个体密度, Dimax为第i个类群的最大密度, g为群落中的类群数, G为各类群所包含的总类群数, Ci/C为相对次数, 即为第i个群落在C个群落出现的次数[13]。
群落相似性:选择Jaccard相似性系数(q)和Gower系数(Sg)描述土壤动物群落的相似性, 公式如下:
Jaccard相似性系数:q=
Gower系数:
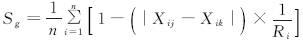
式中, a为A模式全部类群数, b为B模式全部类群数, c为A、B两模式共有的类群数。n为两个群落相比较的类群数; Xi为第i类群的个体数; j和k代表不同的群落; Ri是第i类群在j和k类群的个体总数。
土壤动物功能团划分:拥有不同生活生长方式, 但在功能上相接近的土壤动物类群, 在一定群落中形成具有相同功能的种团, 根据不同的种团功能, 一般划分为腐食性、捕食性和植食性3类功能团[14, 15, 16]。
采用SPSS 17.0对所测数据进行统计分析, 对不同土层深度土壤动物个体密度及类群数进行单因素方差分析, 并用LSD及Duncan法进行多重比较; Excel 2007制图。
本次调查共获得土壤动物303只, 分属3门7纲17目(表1)。在全部土壤动物中, 以线虫纲、膜翅目及蜱螨目为优势类群, 其个数占总捕量的61.06%; 常见类群为鞘翅目幼虫(Coleoptera)、后孔寡毛目(Oligochaetaopisthopora)、弹尾目(Collembola)、倍足纲(Diplopoda)、蜘蛛目(Araneae)、原尾目(Protura)、直翅目(Orthoptera)、等足目(Isopoda)和地蜈蚣目(Geophilomorpha), 占到总捕量的35.64%; 其余5个类群为稀有类群, 共占3.3%。
| 表1 不同模式下土壤动物类群及个体数组成 Table 1 Groups and individuals of soil animals in four compound planting patterns |
模式Ⅰ 包含15个类群, 其中以线虫纲、膜翅目及蜱螨目为优势类群, 共占57.95%, 余下各类群为常见类群; 在模式Ⅱ 的12个类群中, 以线虫纲、膜翅目、鞘翅目幼虫和后孔寡毛目为优势类群, 占到其群落个体总数的72.62%, 其余均为常见类群; 在模式Ⅲ 中的11个类群中, 优势类群为线虫纲、膜翅目和鞘翅目幼虫, 占到模式Ⅲ 总个体数的71.08%, 其余均为常见类群; 模式Ⅳ 共含9个类群, 其中以线虫纲、膜翅目、蜱螨目、鞘翅目幼虫及后孔寡毛目为优势类群, 占到87.5%, 剩余为常见类群。在每个群落内, 由于基数较小, 计算后显示均无稀有类群, 但在全部土壤动物中显示, 鞘翅目成虫(Coleoptera)、鳞翅目幼虫(Lepidoptera)、缨翅目(Thysanoptera)、石蜈蚣目(Lithobiomorpha)及等翅目(Isoptera)为稀有类群。在4个群落中, 线虫纲和膜翅目均为优势类群, 占全部土壤动物个数的49.83%, 且线虫纲作为第一优势类群, 占到总体个数的30.36%。土壤动物类群数和个体数量均呈现出模式Ⅰ > 模式Ⅱ > 模式Ⅲ > 模式Ⅳ 。方差分析可知, 3种林草模式下的土壤动物个体数量都显著高于纯林模式(P< 0.05), 土壤动物类群数较后者也有明显的增长, 表明林草模式下的土壤动物丰富度优于纯林模式, 林+草的复合结构能有效增长土壤动物的数量及种类, 尤其以核桃+苜蓿(模式Ⅰ )达到最大。
土壤动物由于自身生理特征以及对环境变化的响应不同决定了其在土壤空间分布的差异。此次调查发现, 模式Ⅰ 、模式Ⅱ 和模式Ⅲ 中的土壤动物的个体分布具有明显的表聚性, 个体密度从上层(0-5 cm)至下层(10-15 cm)依次减少, 但与3种林草模式不同的是, 在模式Ⅳ 中土壤动物个体密度表现为中层> 下层 > 上层, 呈现出“ 中间大, 两头小” 的纺锤型结构分布(图1)。方差分析表明, 模式Ⅱ 中土壤动物个体密度在各土层间没有显著差异(P> 0.05), 但模式Ⅰ 、模式Ⅲ 和模式Ⅳ 土壤动物个体密度在最密集土层与其它土层中差异显著(P< 0.05), 进一步LSD多重比较表明, 模式Ⅰ 和模式Ⅲ 上层的土壤动物个体密度与中层和下层之间差异均显著(P< 0.05), 两个模式中层和下层的土壤动物个体密度相互差异均不显著(P> 0.05); 模式Ⅳ 中层的土壤动物个体密度与上层和下层之间差异显著(P< 0.05), 而上、下层之间没有显著差异(P> 0.05)。
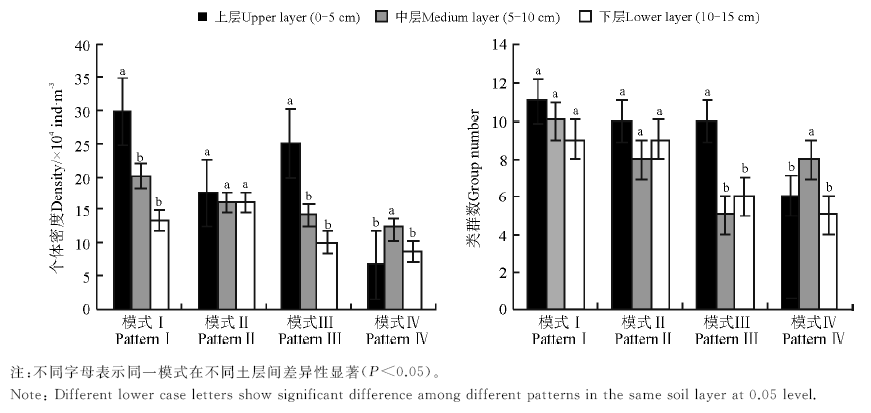 | 图1 不同种植模式下土壤动物群落的垂直结构分布Fig.1 Vertical distribution of soil animals in different soil layers |
除模式Ⅳ 外, 模式Ⅰ 、模式Ⅱ 和模式Ⅲ 的上层土壤动物类群数均大于中层和下层, 但比较中层和下层类群数可以发现, 4种模式下两层土壤动物类群数较接近, 甚至在模式Ⅲ 和模式Ⅳ 中表现出非完全逆向性分布的现象(图1)。经方差分析表明, 模式Ⅰ 及模式Ⅱ 各土层间的土壤动物类群数差异均不显著(P> 0.05), 但模式Ⅲ 0-5 cm土层与其它土层间的土壤动物类群数差异显著(P< 0.05), 中层类群数与下层差异不显著(P> 0.05)。模式Ⅳ 的中层土壤动物类群数显著高于上、下层(P< 0.05), 而上层类群数与下层之间差异不显著(P> 0.05), 在模式Ⅳ 的中层无论是类群数及个体密度均占优势。
Shannon-Wiener多样性指数(H)值排序为模式Ⅰ > 模式Ⅱ > 模式Ⅳ > 模式Ⅲ , 可以看出, 模式Ⅰ 拥有相对丰富的物种, 模式Ⅲ 则相对较寡; Pielou均匀度指数(J)值排序为模式Ⅳ > 模式Ⅱ > 模式Ⅰ > 模式Ⅲ , 在模式Ⅲ 中均匀度指数J最小, 但Simpson优势度指数(C)却最大, 然后依次为模式Ⅰ 、模式Ⅳ 及模式Ⅱ , 综合均匀度指数(J)和优势度指数(C)可知, 模式Ⅲ 群落中土壤动物分布较不均匀, 部分类群优势明显(表2)。模式Ⅳ 土壤动物个体数与其它3种模式差异较大, 总数较小, 分布相对较均匀, 模式Ⅱ 中均匀度指数(J)仅次于模式Ⅳ 的, 且优势度指数C最小, 说明模式Ⅱ 中土壤动物分布较均匀, 无明显的优势类群, 模式Ⅰ 稍次于模式Ⅱ 。密度-类群指数(DG)值排序为模式Ⅰ > 模式Ⅱ > 模式Ⅲ > 模式Ⅳ , 即3种林草模式较纯林模式有更丰富的土壤动物类群及数量。
| 表2 不同种植模式下土壤动物群落多样性指数 Table 2 The diversity indices of soil animals in four compound planting patterns |
选择Jaccard相似性系数(q)研究各模式群落间类群的相似性, 依系数值划分, 在0~0.24为极不相似, 0.25~0.49为中等不相似, 0.50~0.74为中等相似, 0.75~1.00为极相似。结果显示(表3), 模式Ⅰ 与模式Ⅱ 、模式Ⅲ 、模式Ⅳ 以及模式Ⅱ 与模式Ⅲ 均为中等相似, 尤其是模式Ⅱ 与模式Ⅲ 间的相似性系数(q)为最大值, 与其它模式相比, 这两种模式之间的类群更具相似性。模式Ⅳ 与模式Ⅱ 和模式Ⅲ 均属于中等不相似, 尤其是模式Ⅳ 与模式Ⅱ 之间的q值最低, 在4种模式中, 两者类群类型相对差异较大。综合来看, 3种林草模式之间的类群具有一定的相似性, 属于中等相似, 仅有核桃+苜蓿(模式Ⅰ )与纯林模式(模式Ⅳ )的类群具有中等相似性, 核桃+菊苣(模式Ⅱ )和核桃+鸭茅(模式Ⅲ )下的类群种类均与纯林模式(模式Ⅳ )之间呈现中等不相似, 但数值都在0.4以上, 差异程度不大。
| 表3 不同模式下土壤动物群落的相似性系数 Table 3 Similarity indice of soil animals in four compound planting patterns |
Gower系数(Sg)反映不同模式群落中土壤动物个体数的相似性。模式Ⅰ 、模式Ⅱ 及模式Ⅲ 相互间的Sg数值均大于其与模式Ⅳ 之间的Sg值, 尤其模式Ⅱ 与模式Ⅲ 的土壤动物个体数最为接近, 模式Ⅰ 与模式Ⅳ 之间的土壤动物个体数差异最大(表3)。总体上, 核桃+菊苣(模式Ⅱ )和核桃+鸭茅(模式Ⅲ )无论是土壤动物类群种类还是个体数与纯林模式(模式Ⅳ )都具有一定差异, 而核桃+苜蓿(模式Ⅰ )虽然在土壤动物个体数上与纯林模式(模式Ⅳ )有明显差异, 但在类群种类上却与纯林模式(模式Ⅳ )最相似。相对于纯林的种植方式, 加入苜蓿草种进行复合经营的, 不仅没有改变原有纯林模式下的土壤动物类群类型, 保持了一定的相似性, 更显著提高了系统中土壤动物的个体数量, 从另一方面表明, 林草复合种植模式在一定程度上有利于土壤动物的生长活动, 增强群落结构的稳定性。
土壤动物数量庞大, 种类繁多, 拥有不同的生活生长方式, 但某些土壤动物类群在功能上相接近, 形成具有相同功能的种团, 在群落生态系统中发挥着重要的作用。本次调查的土壤动物主要划分为腐食性、捕食性和植食性3类功能团, 腐食性同功能团对土壤形成及生态系统内物质能量流通影响巨大, 通常作为衡量土壤肥高低与生态系统稳定性强弱的生态指标之一[16, 17]; 捕食性同功能团通过捕食行为可在一定程度上有效控制生态系统内的种群数量; 植食性同功能团通常为一些危害林地植被根茎的害虫。
全部模式土壤动物同功能种团组成均表现出腐食性> 捕食性> 植食性(除模式Ⅳ 中捕食性动物类群数与植食性相同), 在4种模式中腐食性动物分别占到72.73%、59.52%、51.81%和64.58%, 捕食性动物分别占20.45%、23.81%、36.14%和20.83%, 植食性动物仅占到其中的6.82%、16.67%、12.05%和14.58%(表4), 综合3种林草模式(模式Ⅰ 、模式Ⅱ 及模式Ⅲ )来看, 腐食性动物在整个林草系统中占到61.57%, 植食性动物占26.67%, 植食性动物占14.51%。可以看到, 腐食性同功能团作为重要的生物肥力指标, 在4种模式中比例均占到绝对优势, 尤其在模式Ⅰ 中比例最大, 达72.73%, 而植食性仅有6.82%, 且模式Ⅰ 拥有最多的土壤动物个体数量, 也说明模式Ⅰ 即核桃+苜蓿拥有相对稳定健康的生态系统和适宜土壤动物生长活动的环境条件。
| 表4 不同模式下土壤动物功能性类群及个体组成 Table 4 The functional groups and individuals of soil animals in four compound planting patterns |
3种林草模式下的土壤动物个体数和类群数皆高于纯林模式, 不同林草模式内的土壤动物个体数量及类群类型也不尽相同, 其中核桃+苜蓿模式生境条件最优越, 土壤动物的个体数和类群数最丰富。此次调查结果表明, 线虫纲、膜翅目和蜱螨目为研究区土壤动物优势类群, 占全部土壤动物个体数的61.06%, 属于亚热带地区森林生态系统典型类群, 这与殷秀琴等[18]对我国土壤动物地理研究的结论一致。调查结果还显示, 线虫为林地优势类群, 与三江平原湿地岛状林[19]和长白山针阔林混交林[2]的研究结果相同, 产生这种情况可能是由于调查时期为夏秋交接时, 降水增多, 同时林下草种还具有一定保水保肥的能力, 保证了微环境的水分水平达到了线虫生长的条件, 使得线虫达到类似湿地林一样的生长优势。
林草模式下的土壤动物空间分布表现出表聚性, 核桃+鸭茅模式土壤动物个体数量垂直分布下降最快, 核桃+苜蓿模式次之, 核桃+菊苣模式变化最缓。纯林模式下的土壤动物个体密度垂直分布呈现出5-10> 10-15> 0-5 cm, 有悖于常论, 根据殷秀琴等[20]对松嫩草原区不同农业生态系统的研究发现, 在气候环境要素中, 地温对土壤动物影响明显。而此次调查在9月展开, 气温依然较高, 纯林模式林下无草种覆盖, 生长初期的核桃林分郁闭不高, 凋落物积累较少, 太阳直射地表, 地表气温较高, 不适宜土壤动物活动, 使得土壤动物不得不往下层活动, 与朱凡等[21]在夏季对节肢动物群落变化的研究结果相同。
Shannon-Wiener多样性指数被广泛作为衡量生态系统群落多样性状况的指标, 然而在本研究发现, 核桃+鸭茅模式无论是土壤动物个体数还是类群数均高于纯林模式下的土壤动物, 但Shannon-Wiener多样性指数值却比纯林模式的值低。因此, 可以看出, Shannon-Wiener多样性指数存在一些局限性, 并不能完整反映出真实的群落多样性状况, 更适用于生态功能较简单的群落。从密度-类群指数结果可以看出, 其规律基本遵循了林地的分布事实, 相比Shannon-Wiener多样性指数, 密度-类群指数综合考虑了个体、类群、均匀度、优势度等问题, 因此, 对于复杂群落的土壤动物多样性程度更具指示作用。
土壤动物的群落分布与地表植被多样性及植被地上和地下的结构关系紧密相关[9, 22]。从密度-类群指数分布可以看出, 不同模式下土壤动物丰富程度依次为核桃+苜蓿> 核桃+菊苣> 核桃+鸭茅> 纯林模式, 3种林草模式皆比纯林模式具有更加丰富的土壤动物群落多样性, 地表植被多样性也比纯林模式丰富, 与土壤动物群落多样性分布一致。在3种林草模式之间比较, 核桃+苜蓿具有更加丰富的群落多样性, 是三者间最适宜的林草模式。在本研究中, 虽然只是根据整体食性将土壤动物群落同功能团粗略地划分成了3个大类, 但有便于研究土壤动物在生态系统中的功能作用, 其数量顺序为腐食性> 捕食性> 植食性。这是由于地下土壤动物的取食途径所决定的, 地下土壤动物不能像地上土壤动物一样直接摄取绿色植物, 主要食物来源是凋落的有机残体, 因而腐食性动物较多, 植食性动物较少。在林草系统中, 草种的保水保肥功能, 使得微环境中营养物质丰富, 水热条件适中, 使得更多土壤动物聚集, 土壤动物间竞争、新陈代谢等作用加剧, 地下腐食性土壤动物拥有更多食物源, 从而形成了良好的生态循环。
通过研究可以看出, 林草系统内土壤动物群落种类丰富, 林下种草不仅能改善土壤动物生存环境条件, 还能促进土壤动物类群及个体的明显增长, 相比纯林模式, 林草模式拥有更加健全稳定的生态系统。不同林草模式内表现也不尽相同, 但关于林草模式下土壤动物与土壤理化性质的关系, 林木与草种之间的作用以及季节动态变化还有待进一步研究。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|