

{kind=link}
{kind=link}
{kind=link}
紫花苜蓿茎秆组织中木质素的分布与沉积模式
[张金青 , 陈奋奇, 汪芳珍, 方强恩, 师尚礼]
, 陈奋奇, 汪芳珍, 方强恩, 师尚礼]
, 陈奋奇, 汪芳珍, 方强恩, 师尚礼]
|
|


第一作者:张金青(1994-),女,甘肃舟曲人,在读本科生,主要从事草种质资源与育种研究。E-mail:18893816768@163.com
以甘农5号紫花苜蓿( Medicago sativa‘Gannong No.5’)为材料,采用组织化学染色方法研究了茎秆中木质素的发生、分布与沉积规律。结果表明,苜蓿茎秆中木质素的分布与生长部位密切相关,在茎秆顶端初生维管组织中仅木质部有木质素分布,随节间下移,木质素开始在初生韧皮纤维、次生木质部和髓射线中大量沉积;茎秆中存在G、S两种木质素,S木质素的发生迟于G木质素;在维管束之间,两种木质素均以“厚角组织处维管束→厚角组织间维管束”的模式沉积,但在维管束内,木质素沉积表现出异质性,G木质素沉积模式为“初生木质部导管→初生韧皮纤维→次生木质部和髓射线”;S木质素沉积模式为“初生韧皮纤维→次生木质部和髓射线→髓薄壁细胞”。分析认为,紫花苜蓿茎秆中木质素特殊的沉积模式可能是其对北温带生长环境的一种适应对策。
This study tested the occurrence, distribution, and deposition of lignin in stem tissue of the alfalfa variety ‘Gannong No.5’ using the histochemical staining method. The research results showed that the distribution of lignin in the stem of alfalfa is closely related to the vascular tissue in different development stages. At the apex of the stem tissue, lignin is only found in the primary xylem. Then, lignin begins to deposit in the primary phloem fibers, secondary xylem, and pith rays, with the internode moving down.Both guaiacyl (G) lignin and syringyl(S) lignin are present in alfalfa stems, and S lignin occurs later than G lignin. Between the vascular bundles, the deposition pattern of the two kinds of lignin is “vascular bundle near the collenchyma→vascular bundles between the collenchyma”. However, the deposition of lignin shows the heterogeneity within the vascular bundle. The deposition pattern of G lignin is “primary xylem vessels→primary phloem fibers→secondary xylem and pith rays”, while the deposition pattern of S lignin is “primary phloem fibers→secondary xylem and pith rays→pith parenchyma”. It is suggested that the special deposition pattern of lignin in the stem of alfalfa is a strategy that is probably appropriate to the growing environment in the north temperate zone.
木质素是由对-香豆醇、芥子醇和松柏醇3种木质醇单体通过多种化学键连接而成的高度异质的大分子肉桂醇脱氢聚合物[1]。这3种木质醇分别形成对-羟基苯基木质素(hydroxyphenyl lignin, H木质素)、愈创木基木质素(guaiacyl lignin, G木质素)和紫丁香基木质素(syringyl lignin, S木质素)[2]。不同植物间及植物不同组织中木质素的种类及其含量不同[3, 4]。最新研究发现, 木质素在植物体内的沉积可增加植物对环境胁迫的抵抗能力[5, 6]。然而, 在细胞壁中木质素却是植物高度抗氧化的一道重要屏障, 是导致牧草消化率降低的主要原因[6]。同时, 在生物燃料生产过程中, 木质素是阻碍纤维素酶降解利用结构性多糖的主要物理性障碍, 严重影响纤维素和半纤维素的工业转化利用[7, 8]。
紫花苜蓿(Medicago sativa)由于其抗逆性强, 适应范围广, 蛋白质含量丰富, 素有“ 牧草之王” 之美称[9, 10]。而且, 紫花苜蓿茎秆中木质纤维素产量高, 可用于生产酒精, 目前被认为是最具有开发潜力的能源植物之一[11, 12]。然而, 茎秆中木质素的沉积严重阻碍着紫花苜蓿的高效利用, Reddy等[13]研究了苜蓿茎秆中木质素的积累对其品质的影响, 发现木质素沉积量的降低, 会显著提高紫花苜蓿在反刍动物瘤胃中的消化率。Chen和Dixon[14]从生物质型(biomass type)苜蓿品种的选育和改良角度研究指出, 降低紫花苜蓿茎秆中木质素的含量, 其糖化效率(saccharification efficiency)最高可增加80%以上。可见, 无论从牧草角度还是从能源作物角度分析, 木质素均是紫花苜蓿高效利用的物理性障碍, 获得低木质素的新型种质是紫花苜蓿育种的重要目标。但是, 关于紫花苜蓿茎秆中木质素的发育机理至今尚未阐明, 木质素在茎秆不同组织中的分布与沉积规律仍不清楚[15]。针对上述分析, 本研究以甘农5号紫花苜蓿为材料, 通过组织化学研究方法, 揭示苜蓿茎秆中木质素的分布与沉积模式, 以期为低木质素新型种质育种研究提供基础资料。
供试材料为甘农5号紫花苜蓿, 种子由甘肃农业大学草业学院提供。该品种是由贺春贵教授等人选育的抗蚜新品种, 具有再生速度快、高产、抗蚜虫等特点, 已于2010年通过国家牧草品种委员会审定为育成品种。
试验在甘肃农业大学兰州牧草试验站进行, 该地区属温带半干旱大陆性气候, 四季分明、光照充足。海拔1 517.3 m, 年平均降水量320 mm, 年日照时数2 474.4 h, 土壤pH约8.2。田间试验小区面积3 m× 3 m, 3次重复。2016年4月上旬播种。播前结合整地施磷肥750 kg· hm-2、氮磷复合肥375 kg· hm-2。条播, 播种行距30 cm, 播量为15 kg· hm-2。播种深度1.5~2.0 cm。田间管理包括间苗、中耕除草、适时灌溉。播种当年分别于7月5日、8月25日两次刈割, 每次留茬高度均为5 cm。
取样:在苜蓿初花期(2017年5月4日), 选取6株长势一致的健康植株, 每株选出最高枝条, 带回实验室, 用Gillette双面刀片从始花节(即茎秆中第1个开花节)自上而下切取每一节间, 并依次升序编号, 对新鲜材料立即进行徒手横切, 切片厚度为20~25 μ m, 所得切片用两种不同的染色方法进行滴染, 制片, 置于日产正置万能显微OLYMPUS-BX61下观测、拍照, 图片用Photoshop CS6软件进行编号整理。
G木质素染色(Wiesner法[16]):将切片置于载玻片上, 滴加数滴2%间苯三酚乙醇溶液(95%乙醇配置)染色3 min, 再滴加3 mol· L-1的盐酸数滴, 染色1 min后封片观察, 组织中G木质素存在部位呈现紫红色。
S木质素染色(Mä ule法[17, 18]):将切片置于载玻片上, 先用0.5%高锰酸钾溶液滴染2 min, 用蒸馏水冲洗切片数次, 然后在3 mol· L-1的盐酸溶液中滴染1 min, 再用蒸馏水洗掉反应后剩余的二氧化锰, 最后用10%的氨水溶液滴染1 min, 封片后观察。组织中S木质素存在部位呈现桔红色, G木质素存在部位呈现黄褐色。
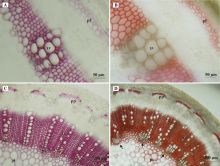
紫花苜蓿茎秆初生维管组织(图1A, B)和次生维管组织(图1C, D)中木质素的类型与分布部位如图1所示。可以看出, 苜蓿茎秆中存在G和S两种木质素, 主要分布在木质部、初生韧皮纤维细胞和髓射线中, 但两种木质素的分布存在差异。图1A显示G木质素的染色结果, 初生木质部导管分子细胞壁染色最深, 其次是髓射线, 初生韧皮纤维细胞染色最浅; 图1B中, 髓射线与初生韧皮纤维显色明显, 而初生木质部导管显黄褐色, 没有染上桔红色, 表明在初生木质部导管分子细胞壁中只有G木质素沉积, 初生韧皮纤维细胞壁中S木质素多于G木质素。在茎秆次生维管结构中, G和S木质素在次生木质部中大量沉积, 此时, 初生木质部中仍未见S木质素, 但在邻近初生木质部的髓薄壁细胞壁中, 出现了S木质素的沉积(图1D, 箭头)。
 | 图1 紫花苜蓿茎秆组织中木质素的类型及分布 A、B为第8节间, 显示茎秆组织的初生结构; C、D为第15节间, 显示茎秆组织的次生结构; A、C显示G木质素的分布(Wiesner染色法); B、D显示S木质素的分布(Mä ule染色法)。pp:初生韧皮部; sx:次生木质部; px:初生木质部; pi:髓; pf:初生韧皮纤维; xv:初生木质导管, 下同。Fig. 1 The type and distribution of lignin in alfalfa stem tissue A, B: The eighth internodes show the primary structure; C, D: The fifteenth internodes show the secondary structure; A and C show the distribution of G lignin(by Wiesner staining method); B and D show the distribution of S lignin(by Mä ule staining method). Abbreviations:pp, Primary phloem; sx, Secondary xylem; px, Primary xylem; pi, Pith; pf, Primary phloem fiber; xv, Primary xylem vessel; similarly for the following figures. |
2.2.1 G木质素 图2显示初花期紫花苜蓿茎秆自上而下第1、第4、第7、第10、第13、第16节间不同发育阶段维管组织中G木质素的染色结果, 可以看出, 在早期初生维管束中, 仅导管分子细胞壁中有木质素沉积(图2A)。随着初生维管束的发育, 初生韧皮纤维中也沉积了少量木质素(图2B、C)。当维管束次生结构形成时, 髓射线和次生木质部中沉积了大量木质素(图2D)。随着茎秆进一步成熟, 木质素积累越来越多, 木质化程度越来越高(图2E、F)。
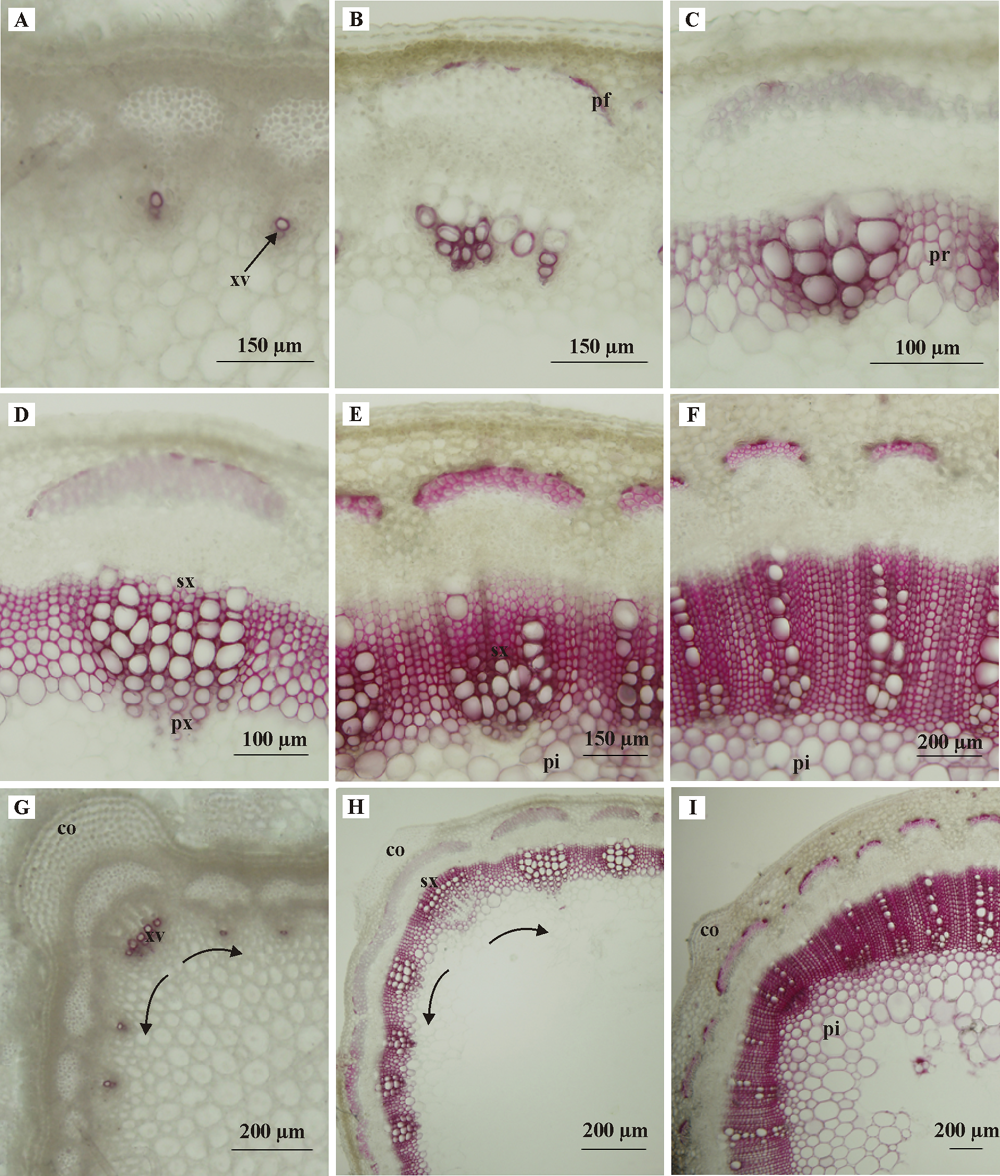 | 图2 紫花苜蓿茎秆不同节位G木质素(Wiesner染色法)的沉积 A、B、C、D、E、F分别是第1、4、7、10、13、16节间, 显示维管束内G木质素的沉积规律; G、H和I分别是第1、7、13节间, 显示维管束间G木质素的沉积规律。pr:髓射线, co:厚角组织。Fig. 2 The deposition of G lignin in different internode of alfalfa stem(by Wiesner staining method) A, B, C, D, E, and F: The 1st, 4th, 7th, 10th, 13th, and 16th internodes show the deposition pattern of G lignin within the vascular bundle; G, H and I: The lst, 7th, and 13th internodes show the deposition pattern of G lignin between the vascular bundles. Abbreviations: pr, Pith ray; co, Collenchyma |
紫花苜蓿茎秆具有4个棱, 从茎的横切面看, 每个棱由径向向外膨大凸出的皮层厚角组织形成(图2-G、H、I)。比较不同发育阶段茎维管结构的变化, 发现茎棱厚角组织处维管束初生结构的发育要早于两侧(图2G、H, 箭头处), 但进入次生生长阶段后, 两侧维管束发育速度加快, 木质化面积不断向髓部扩大, 此时茎棱处维管束发育相对滞后。木质素的沉积也显示出与之相对应的特点。
上述分析表明, G木质素在苜蓿茎秆组织中的沉积遵循以下规律:在维管束内, 木质素以“ 初生木质部导管→ 初生韧皮纤维→ 次生木质部和髓射线” 的模式沉积。在维管束之间, 木质素最先在厚角组织处维管束初生结构中沉积, 随之再向两侧维管束延伸; 在次生生长阶段, 两侧维管束中木质素沉积速度加快, 沉积面积不断向髓中心扩大。
2.2.2 S木质素 S木质素在维管束内的沉积模式与G木质素相似, 也是随节间下移木质素沉积增多, 木质化程度升高(图3)。在初生维管束早期没有S木质素的沉积(图3A)。随着初生维管束的发育, 初生韧皮纤维细胞壁中开始有少量S木质素的积累(图3B)。当次生维管束逐步发育形成时, S木质素在次生木质部和髓射线中大量出现(图3C、D、E)。在茎秆组织进一步成熟后, 髓薄壁细胞壁中也出现了大量的S木质素(图3F)。
 | 图3 紫花苜蓿茎秆不同节位S木质素(Mä ule染色法)的沉积 A、B、C、D、E、F分别是第1、4、7、10、13、16节间, 显示维管束内S木质素的沉积规律; 图G、H、I分别是第4、10、16节间, 显示维管束间S木质素的沉积规律。Fig. 3 The deposition of S lignin in different internode of alfalfa stem(by Mä ule staining method) A, B, C, D, E, and F: The 1st, 4th, 5th, 10th, 13th, and 16th internodes show the deposition pattern of S lignin within the vascular bundle; G, H and I: The 4th, 10th, and 16th internodes show the deposition pattern of S lignin between the vascular bundles. |
S木质素在维管束之间的沉积规律与G木质素相同, 即在维管束初生结构中, S木质素首先在厚角组织处的初生韧皮纤维中沉积, 然后在两侧积累(图3G, 箭头)。随着维管束次生结构的发育, 两侧维管束中的木质化速度加快, 而厚角组织处维管束的木质化速度相对较慢(图3I)。
上述分析表明, 在维管束内, S木质素以“ 初生韧皮纤维→ 次生木质部和髓射线→ 髓薄壁细胞” 的模式沉积。在维管束之间, S木质素以“ 厚角组织处维管束→ 厚角组织间维管束” 的模式沉积。
对比观察图2、图3可以发现, 虽然两种木质素单体的沉积模式相似, 但S木质素的沉积迟于G木质素; 初生木质部导管中未发现S木质素, 在次生生长阶段, 髓薄壁细胞壁中出现了S木质素的沉积。
木质素作为细胞壁次生壁的三大主要成分之一, 在维管植物结构的坚硬化、水肥运输和抗病性方面具有重要作用[19]。本研究发现, 紫花苜蓿茎秆初生木质部导管中只有G木质素沉积, 而初生韧皮纤维细胞中S木质素占优势(图1A、B)。Menden等[20]研究指出植物在受到病原物侵染时会选择性增加S木质素单体的含量。木质素具有疏水性, 导管是木质部中最主要的输导组织[21], 因此木质部中G木质素的沉积可增强其对水分的输导能力[16]。由此分析, 苜蓿茎秆初生韧皮纤维中S木质素的大量沉积可能与抗病有关, G木质素的沉积主要与组织结构坚硬化和水肥运输有关。
G木质素单体易于和其他基团结合形成稳定的化合物而较S木质素单体更难降解[22]。因此普遍认为, 通过降低牧草中G/S的比例可以改善牧草的品质, 提高牧草的消化率[23], 但是这一观点目前仍存在争议[22]。Reddy等[13]研究指出, 降低木质素的含量可以提高反刍动物对苜蓿的消化率, 但苜蓿中G/S比值的改变并没有对消化率产生明显影响。G、S木质素对消化率的影响机理目前尚不清楚, 因此, 为了提高牧草消化率, 简单降低G/S值可能并不合理。另外, 从生物燃料生产角度讲, 现有研究结果表明, 降低紫花苜蓿中S木质素单体的含量, 可显著增加生物酒精的产量[23]。在杨属(Populus)植物中, G/S值的升高与酒精和糖产量均呈正相关[24, 25]。但是, 就目前研究结果看, 木质素含量及其组成与生物质能的转化效率的关系仍然存在较大争议[12]。在苜蓿中, S木质素含量降低可增加酒精产量, 但G/S值的改变对糖化效率的影响并不明显[14]。Pedersen等[26]研究指出, 降低高粱(Sorghum bicolor)、柳枝稷(Panicum virgatum)、紫花苜蓿等能源作物木质素的含量, 并不能显著提高生物燃料产率。Fu等[3]研究了柳枝稷中木质素组成与生物质能的转化关系, 结果表明, 随着G/S值的升高, 酒精产量显著增加, 但Sarath等[27]的研究却取得了截然相反的结果, 认为G/S值的改变与酒精产量无关。木质素的分布与沉积随植物种类及其组织部位不同而存在差异[28]。本研究结果也表明, 苜蓿茎秆中两种木质素的沉积量与发育阶段和分布部位相关。可见, 植物中木质素的发生发育过程与生物功能复杂多变, 要完全揭示木质素的作用机能, 还需要从分子水平进一步深入研究。
Vanholme等[29]从生物适应性进化角度分析, 认为木质素的沉积是植物对于陆生生活环境的适应, 主要为植物的直立生长提供支撑作用。本研究结果显示, 苜蓿同一节位茎秆组织中维管束之间木质素的发生与沉积并不同步, 在茎维管束初生生长时期茎棱厚角组织处维管束内木质素的沉积早于其两侧维管束(图2G、H, 3G)。目前普遍认为, 紫花苜蓿起源于“ 近东中心” — — 小亚细亚、外高加素、伊朗和土库曼的高地, 这些地区的特点是春季迟临, 夏季短促[30], 木质素在维管束中这种独特的发生特性可能有利于其返青期再生枝的快速直立生长, 这可能也是苜蓿对北温带大陆性气候短暂生长季的一种适应对策。
木质素沉积的异质性是指随着植物种类、个体、器官组织、细胞类型以及环境条件的不同, 其组成、含量和结构也不完全相同, 甚至变化很大, 这是木质素生物合成过程中不同类型酶时空表达的结果[31, 32, 33]。在拟南芥(Arabidopsis thaliana)木质素沉积突变体中, 木质素异位沉积于髓薄壁细胞中, 这种异位沉积在幼茎和成熟茎中都存在[5], 但本研究发现, 在紫花苜蓿幼茎初生木质部中只有G木质素沉积, 而在成熟茎秆髓薄壁细胞中存在S木质素, 这与Saito等[34]在鸡爪槭(Acer micranthum)中的研究结果一致。截至目前, 只能得到木质素的结构模型, 仍未能获得完整的天然木质素单体[35], 紫花苜蓿茎秆中木质素的异质性沉积也许能为天然木质素单体提取提供一条可行的途径。
(责任编辑 张瑾)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|