

{kind=link}
{kind=link}
{kind=link}
{kind=link}
黄帚橐吾基株克隆生长对不同光照的响应
[王满堂1  , 赵全科
, 赵全科1 , 程栋梁2 ]
, 赵全科]
|
|


第一作者:王满堂(1980-),男,山东枣庄人,副教授,博士,主要从事植物生态学科研工作。E-mail:wangmantang@aliyun.com
为了解青藏高原高寒草甸克隆植物黄帚橐吾( Ligularia virgaurea)的生活史对策,通过田间盆栽控制试验,模拟青藏高原东部高寒草甸上由于过度放牧产生的裸地和低、矮的退化草甸的光强,研究黄帚槖吾基株的相对克隆生长速度和克隆性状对不同光照的响应,以期填补有关黄帚槖吾克隆生长领域研究的不足,并为该类毒杂草的防治提供理论依据。结果表明,在自然光照条件下,黄帚橐吾根状茎长度与时间无线性关系;在遮阴环境下,黄帚橐吾根状茎长度与时间之间存在显著的线性关系( P<0.05),根状茎长度随着时间延长而显著增长。光照不仅可以直接影响单位长度根状茎生物量,还可以通过影响基株大小从而间接影响单位长度根状茎生物量。黄帚橐吾根状茎的长度与单位长度根状茎生物量之间不存在权衡。光照对黄帚橐吾克隆生长相对速率有显著影响( P<0.05),遮阴条件下黄帚橐吾克隆相对生长速率显著大于自然光照条件下的生长速率。因此,在遮阴下根状茎主要功能是觅食,自然光照下根状茎主要功能用于贮藏养料、固定作用。
Understanding the life history strategies of Ligularia virgaurea in the alpine meadow of the Qinghai-Tibet plateau is important to preventing and controlling this weed. We studied the response to light intensity of clonal growth and traits of L. virgaurea genets in field pots experiments to fill the gap in research and provide a theoretical basis for the prevention and control of this weed. Two light intensity levels were used to simulate the light intensities in bare soil and in low, open vegetation of disturbed meadows on the eastern Qinghai-Tibet plateau. The results showed that there was not a linear relationship between the length of rhizomes in L. virgaurea and the time under full natural irradiance. In shaded conditions, there was a significant positive linear relationship between the length of rhizomes in L. virgaurea and time ( P<0.05). Light intensity directly affected the dry mass per unit rhizome length. It also indirectly affected the dry mass per unit rhizome length by increasing the dry mass of the genets. There was no trade-off between the length of rhizomes and the dry mass per unit rhizome length in L. virgaurea. Light intensity had a significant impact on relative clonal growth rates of L. virgaurea ( P<0.05). The relative clonal growth rates under shaded conditions were significantly higher than those under full natural irradiance. Therefore, the main function of rhizomes is foraging in shaded conditions, but, under the full natural irradiance, their primary role is storage and anchoring.
青藏高原东北缘的高寒草甸是我国重要的天然牧场。由于过度放牧、乱采滥挖等人类活动的干扰和全球气候变化等因素的影响, 该地区高寒草甸生态系统退化日趋严重。作为青藏高原高寒草甸退化的指示性植物— — 黄帚橐吾(Ligularia virgaurea)大量扩散和蔓延[1]。20世纪70年代末, 黄帚橐吾盖度为5%~20%, 而1992-1996年上升到40%~60%[2]; 2002年对青藏高原东北缘6个不同海拔梯度的样点调查显示, 均有大面积的黄帚橐吾斑块或与牧草混生群落[3, 4]。2006年, 笔者所在课题组对该区域3种不同生境样点调查显示, 滩地密度为25株· m-2, 坡地为33株· m-2, 沙地为24株· m-2。因此, 研究青藏高原高寒草甸这一典型克隆植物— — 黄帚橐吾的生活史对策具有重要的应用价值。
克隆植物的克隆生长特性是生态和进化研究的主题和热点, 目前许多研究集中于克隆植物如匍匐茎和根茎类草本植物的生理整合、觅食行为和克隆生长策略[5, 6, 7, 8]。对于一些多年生的克隆草本植物, 其生活史的适合度受到很多因子的影响, 如繁殖方式(有性繁殖和克隆繁殖)、时间(当前克隆繁殖与未来有性繁殖)等。在每次繁殖时, 克隆植物通常要把同一资源分配给生长、存活、无性繁殖和有性繁殖等不同生活史过程。由于资源的限制, 克隆植物生长与繁殖、有性繁殖和无性繁殖对有限的资源均存在着权衡关系。以往这种权衡关系的研究多以克隆植物分株为研究对象[9, 10], 很少以克隆植物基株为研究对象[11]。
光是青藏高原高寒草甸重要的环境因子之一, 是影响植物生长和生物量分配的重要因素之一[12, 13]。研究发现, 光强变化对植物产生自然选择压力, 从而影响叶片生理生态特征和植物个体生物量在不同器官间的分配[14]。此外, 光照强度还影响克隆构件特征, 如在低光照条件下根状茎会较短[15], 但也有研究发现根状茎长度在低光照下增长[16]。黄帚橐吾的根状茎不仅具有觅食功能, 还具有贮藏养料功能, 这两大功能间是否存在权衡及这种关系对光强变化的响应等尚待研究。
为了揭示青藏高原高寒草甸典型克隆植物黄帚橐吾的生活史对策, 于2004年10月至2006年10月通过野外田间盆栽控制试验, 主要解决以下问题:1)光强对黄帚槖吾基株的克隆相对生长速率的影响; 2)光强对黄帚橐吾克隆性状如根状茎长度、单位长度根状茎生物量的影响。
研究地点设在甘肃省甘南藏族自治州合作市(102° 53' E, 34° 57' N, 海拔2 900 m)。该地区年平均气温2.0 ℃, 最冷的冬季3 个月平均气温-8.9 ℃, 最热的夏季3 个月平均气温为11.5 ℃; 年均降水量660 mm[4, 17]。
黄帚橐吾为菊科(Compositae)橐吾属(Ligularia)多年生草本, 灰绿色, 根肉质, 簇生[4, 17]。广泛分布于青藏高原东部、东南部, 主要分布区为甘肃省南部、青海省东南部、四川省和云南省西北部[3]。主要生活在海拔2 600~4 700 m、水分适中的河滩、沼泽草甸、阴坡湿地及灌丛中。其营养生长期地上部分无茎, 仅有1~10片簇生叶。黄帚橐吾既能进行有性繁殖, 又能进行克隆繁殖, 是典型的游击型(guerilla)克隆生长植物[4, 17]。黄帚橐吾实生苗1 龄株无克隆繁殖和有性繁殖[12]。
2004 年9月中旬至10月上旬, 在甘南州玛曲县(35° 58' N, 101° 53' E)城郊随机采集黄帚橐吾种子。为保证所采种子已经成熟, 在种子开始扩散时采集。去除败育、虫害的种子后, 把所有种子混合在一起, 装入牛皮纸信封, 室温保存。
根据青藏高原东部高寒草甸上由于过度放牧产生的裸地和低、矮的退化草甸的光强[18], 本研究设自然光照(100%透光)、遮阴(75%遮阴)两个处理, 遮阴处理采用75%的标准遮阴网搭遮阴棚, 依据照度计测量调整遮阴棚高度。试验于2005年4月播种在高25 cm、直径30 cm花盆里并编号, 每盆播40粒。花盆的土壤于2004年取自原生境, 且经薄膜覆盖数月消除土壤中的种子, 用孔径为2 mm的筛子筛两遍, 充分混匀。花盆表面撒一层薄沙, 有利保水。每个处理27个重复, 每个花盆随机排放。不同处理种子出苗率约为65%。每个光照处理保证每盆有20 株幼苗。
2006 年分别在返青后50、100、150 d(第2生长季末)取样, 每次从不同光照梯度随机取95株苗。部分2龄株的黄帚橐吾实生苗进行克隆繁殖[12], 自然光照下每次取样发生克隆繁殖的株数依次为39、36、34, 遮阴下每次取样发生克隆繁殖的株数依次为19、25、19。
每次取样后, 将产生克隆繁殖的个体分为叶片、叶柄、根、根状茎、克隆分株等部分, 滤纸吸干表面水分, 并测量根长、根状茎长、叶柄长。用扫描仪测出叶面积, 然后放入80 ℃烘箱中烘干48 h后用精度为万分之一的电子天平测其重量。
黄帚橐吾克隆结构(clonal structure)是指根状茎及克隆分株。单位长度根状茎生物量计算公式如下:
单位长度根状茎生物量=根状茎干重/根状茎长度。
采用单因子方差分析研究光照对黄帚橐吾根状茎长度的影响。协方差分析研究光照对黄帚橐吾单位长度根状茎生物量的影响, 基株干重为协变量, 光照为固定因素。
黄帚橐吾根状茎长度、克隆结构干重依次与时间进行线性回归分析, 使用最小二乘法(ordinary least squares regression, OLS)计算, 其斜率分别表示根状茎长度的相对增长速率、黄帚橐吾克隆生长的相对速率。使用标准主轴回归方法(standardized major axis, SMA)进行单位长度根状茎生物量与基株大小、单位长度根状茎生物量与根状茎长度的回归分析。
黄帚橐吾的所有变量均经过自然对数转换。所有回归分析使用软件Standard Major Axis Tests and Routines(SMATR)分析[19], SMATR软件还被用来检测不同光照间直线斜率、截距是否存在显著差异[20]。所有的方差分析均使用SPSS 16.0软件, 作图由Origin 9.0软件完成。
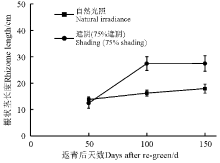
光照对黄帚橐吾根状茎长度的影响与生长季关系密切。在返青后50 d, 光照对黄帚橐吾根状茎长度没有显著影响(F=1.968, P=0.166); 在返青后100和150 d, 光照对黄帚橐吾根状茎长度产生了显著影响(100 d, F=8.980, P=0.004; 150 d, F=7.678, P=0.008)(图1)。自然光照下, 黄帚橐吾根状茎与时间之间没有线性关系; 遮阴环境下, 黄帚橐吾根状茎长度与时间之间存在显著的线性关系, 且随着时间增加而显著增长(图1, 表1)。
| 表1 不同光照处理下根状茎长度与生长时间的回归 Table 1 Parameter estimation for the relationship between Ligularia virgaurea rhizome length and growing time |
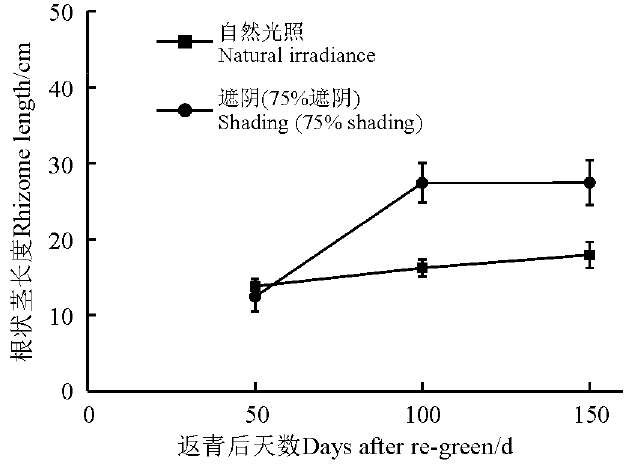 | 图1 第2生长季不同光照条件下黄帚橐吾根状茎长度Fig. 1 Rhizome length under different light conditions in the second growing season |
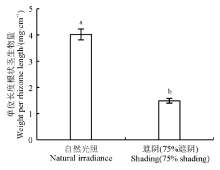
黄帚橐吾的基株干重与单位长度根状茎生物量之间存在显著的正相关关系(图2)。不同光照条件下, 两条直线的斜率没有显著差异(P> 0.05)而截距存在显著差异(P< 0.001), 表明单位长度根状茎生物量随着基株大小增加的速率一样, 但增加的绝对值不一样。经协方差分析, 发现光照对黄帚橐吾单位长度根状茎生物量产生显著影响(F=78.93, P< 0.001)(图3)。光照不仅可以直接影响单位长度根状茎生物量, 还可以通过对基株大小的影响间接影响单位长度根状茎生物量。遮阴环境下, 整个生长季黄帚橐吾根状茎的长度持续增加, 而单位长度根状茎生物量与根状茎长度呈显著的正相关关系(P< 0.05), 表明根状茎长度与单位长度根状茎生物不存在权衡。自然光照下, 虽然黄帚橐吾根状茎长度增加到一定长度就停止, 单位长度根状茎生物量与基株大小、根状茎长度均呈显著的正相关关系, 因而根状茎长度与单位长度根状茎生物量也不存在权衡关系。综上所述, 黄帚橐吾根状茎的长度与单位长度根状茎生物量之间不存在权衡。
 | 图2 第2生长季不同光照条件下黄帚橐吾单位长度根状茎生物量与基株大小的回归Fig. 2 Regression of genet weight and mean weight per Ligularia virgaurea rhizome length under different light conditions in the second growing season |
 | 图3 第2生长季不同光照条件下黄帚橐吾单位长度根状茎生物量 不同小写字母表示不同处理间差异显著(P< 0.05)。Fig. 3 Mean weight per Ligularia virgaurea rhizome length under different light conditions in the second growing season Values with different lowercase letters show significant differences at the 0.05 level. |
黄帚橐吾克隆繁殖结构的干重与时间之间存在显著的线性关系(图4, 表2)。因而, 自然光照条件下和遮阴条件下黄帚橐吾克隆生长相对速率分别为0.03和0.04 g· d-1。不同光照下, 这两条直线的斜率具有显著差异, 表明光照对克隆生长相对速率存在显著影响。遮阴下黄帚橐吾克隆相对生长速率显著大于自然光照条件。
| 表2 不同光照处理下克隆繁殖结构与生长时间的回归 Table 2 Parameter estimates for the relationship between of L. virgaurea clonal structure weight and growing time under different light conditions in the second growing season |
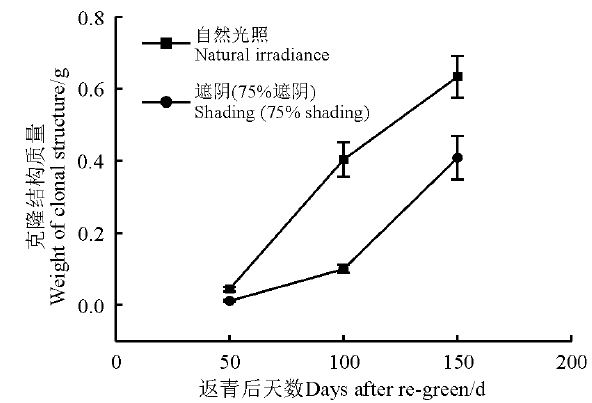 | 图4 第2生长季, 不同光照下黄帚橐吾克隆部分干重Fig. 4 Dry weight accumulation of clonal structures under different light conditions in the second growing season |
自然光照下, 黄帚橐吾根状茎长度的增长速率显著小于遮阴下根状茎的增长速率, 并且自然光照条件下, 根状茎长到一定长度就停止生长, 根状茎数量也多于遮阴下根状茎的数量, 根状茎的主要功能为固定作用, 而在遮阴环境下根状茎主要起到觅食作用。因此, 克隆植物能够辨别生境的优劣。该研究结果与Ikegami等[21]研究的莎草(Scirpus olneyi)根状茎长度受到光照影响的结果一致。
克隆植物根状茎具有两大功能, 一是在异质性环境中, 通过形态可塑性, 把无性克隆分株放置在有利的环境, 从而实现觅食行为[22]。本研究的遮阴处理模拟了黄帚橐吾在退化草甸群落中获得的光照强度, 而自然光照处理模拟了在过度放牧情况下裸地斑块的光强, 研究发现, 自然光照下根状茎长度较短, 黄帚橐吾占据生境的能力增强, 表明在过度放牧情况下, 裸露的斑块适宜黄帚橐吾的拓殖。因此, 过度放牧促进了黄帚橐吾种群扩大的机会, 并对高寒草甸造成破坏。
克隆植物根状茎的另外一个功能是贮藏, 可用于更新的芽和光合产物[23]。根状茎的此功能可以为黄帚槖吾克隆分株的成苗和生长以及休眠芽的长期存活提供物质保证, 从而确保克隆生长的顺利进行, 提高黄帚橐吾基株的适合度。当根状茎作为储存器官时, 基株首先尽可能地满足叶和根的生长, 在资源供应允许的情况下才开始根状茎的生长, 进行克隆繁殖[15]。从对黄帚橐吾基株大小与单位长度根状茎生物量的回归分析可知二者存在正相关关系, 并且光照对二者关系没有显著的影响。黄帚橐吾的单位长度根状茎生物量受基株大小的影响, 基株越大, 越能够制造更多的光合产物转移到根部, 根状茎在光合同化物上接受基株的支持。光照不仅通过对基株大小的影响从而间接影响单位长度根状茎生物量, 而且还直接影响单位长度根状茎生物量, 光照越强其生物量越大。根状茎的两大功能之间不存在权衡。
自然光照下, 黄帚橐吾克隆生长相对速率小于遮阴下的生长速率, 而单位长度根状茎生物量大于遮阴下的单位长度根状茎生物量, 且根状茎长度、根状茎数量均小于遮阴下的, 因而可以推测, 在遮阴下根状茎主要功能是用于逃离不利的生境, 自然光照下根状茎主要功能用于贮藏养料、固定作用。因此, 黄帚橐吾根状茎的生理生态特征和功能对环境具有响应。
(责任编辑 武艳培)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|