

{kind=link}
{kind=link}
{kind=link}
艾丁湖盐角草种群动态生命表及其对温度变化的响应
引用本文
王锋, 黄俊华, 杨文英, 包灵, 仲亚婷, 刘灿. 艾丁湖盐角草种群动态生命表及其对温度变化的响应. 草业科学, 2017,34(5):1064-1071
Wang Feng, Huang Jun-hua, Yang Wen-ying, Bao Ling, Zhong Ya-ting, Liu Can. Dynamic life table and response to temperature change of Salicornia europaea in Ayding Lake . Pratacultural Science,2017,34(5): 1064-1071
Permissions
Wang Feng, Huang Jun-hua, Yang Wen-ying, Bao Ling, Zhong Ya-ting, Liu Can. Dynamic life table and response to temperature change of Salicornia europaea in Ayding Lake . Pratacultural Science,2017,34(5): 1064-1071
Copyright©2017, 《草业科学》编辑部
《草业科学》编辑部
艾丁湖盐角草种群动态生命表及其对温度变化的响应
第一作者:王锋(1991-),男,甘肃天水人,在读硕士生,主要从事野生动植物保护与利用方面的研究。E-mail:791114875@qq.com
摘要
艾丁湖地处亚洲中部极端干旱区,夏季酷热,地势闭塞,环境条件独特。本研究在该地区选择3个具有不同水分特征和盐角草( Salicornia europaea)种群特征的样地(A样地:地形偏高,4月之后地表无积水,盐角草密度相对较低;B样地:位于地表径流覆盖范围内,地表长期有积水,种群密度相对中等;C样地:地表早期有积水,种群密度相对最大,盖度最高)进行调查,根据盐角草的生长情况,从3月至10月每月调查1~2次,共10次,每次统计样方内盐角草的株数。利用动态生命表以及相关性分析的方法来研究该地区盐角草种群变化及其与环境温度之间的关系。结果表明,艾丁湖盐角草种群密度在5月前基本呈增长状态,5月末至6月由于温度升高、水分下降和营养物质减少引发了种群自疏,出现第1次死亡率高峰期;7月至8月,A、B两个种群出现第2次死亡率高峰期,主要原因是连续长达34 d的40 ℃以上的极端高温;9月A种群出现第3次死亡率高峰期,主要原因可能是高温以及高温加速土壤水分蒸发引起的土壤返盐、水分丧失,从而导致的高温胁迫和盐胁迫。相关性分析表明,A、B种群死亡率与高温显著相关( P<0.05)。C种群仅有一次死亡率高峰期,且与温度相关不显著( P>0.05),与A、B种群差异较大的原因可能是由于C样地水分条件好、种群密度高缓解了高温的胁迫作用。A、B两个种群存活曲线属于Deevy-Ⅲ型,即初期死亡率较高,9月之后趋于稳定;C种群属于Deevey-Ⅱ型存活曲线,呈对角线型,即各个龄级存活率基本一致。
关键词:
种群动态; 高温胁迫; 盐胁迫; 存活曲线; 极端环境
中图分类号:S812.29
文献标志码:A
文章编号:1001-0629(2017)05-1064-08
doi: 10.11829/j.issn.1001-0629.2016-0424
Dynamic life table and response to temperature change of Salicornia europaea in Ayding Lake
Abstract
Ayding Lake, located in the extreme arid region in Middle Asia, has a unique environment condition. Occluded terrain leads to high temperatures in summer. Currently, no studies have shown how this area is affected by Salicornia europaea. In this study, 3 sample plots with different water and population characteristics were selected and named plots A, B, and C. Plot A was a high terrain, with no water on the surface after April and low plant density. The surface of plot B was covered with water for a long period, and it had moderate plant density. The surface of plot C was covered with water during the early months of the year, and it had the highest plant density. We set 3 quadrats of 0.5 m×0.5 m in each plot. Each quadrat was investigated 1~2 times a month, and the plant density was counted 10 times. By using dynamic life table and correlation analysis, we studied the relationship between population change and temperature. The results showed that S. europaea populations grew before May. In all plots, the high mortality period for S. europaea populations was from May to June because of self-thinning triggered by increase in temperature and reductions in moisture and nutrients. The S. europaea populations of plots A and B showed a second high mortality period from July to August because of the extreme temperature of 43 ℃ for 43 days. The S. europaea population in plot A showed a third high mortality period in September because of high temperature and salt stresses caused by evaporation of soil moisture resulting in increased salt concentration. The plant mortality noted in plots A and B was significantly correlated with temperature. S. europaea population in plot C had only one period of high mortality, but this was not significantly correlated with temperature. This is because compared to plots A and B, plot C had more moisture and higher plant population density, which can relieve the stress of high temperature. The survival curves of plant populations in plots A and B belong to Deevey type Ⅲ, which show high mortality rates at the beginning of the year, but tend to become steady with time. The survival curve of plant population in plot C belongs to Deevey type Ⅱ, which shows the same mortality rate at all times in a year.
Keyword:
population change; high-temperature stress; salt stress; survival curve; extreme environment
在干旱和半干旱地区, 由于水分的缺失易导致土壤盐渍化[1], 进而限制农业的发展[2], 然这种恶劣的自然条件使丰富的抗盐品种孕育而生[1]。盐角草(Salicornia europaea)是藜科(Chenopodiaceae)盐角草属(Salicornia)一年生双子叶草本植物, 也叫“ 欧洲海蓬子” [3], 较其近缘种海蓬子而言, 适生范围更广, 广泛分布于中亚、北美以及日本等地区[4]; 在我国主要分布于甘肃西部、青海、新疆、西藏等地[5]。
盐角草是世界上发现的最抗盐的高等植物之一[6], 具有特别强的摄盐能力, 它在生长过程中能淡化周围盐渍土的含盐量, 因此对于盐渍土的改良具有重要的意义[7]。另外, 盐角草的种子含油量高, 有望开发成油料作物[4]; 植株体蛋白质含量丰富, 可以作为饲料; 盐角草也是主要的耐盐基因的来源, 对于提高农作物的耐盐能力和生态适应能力有重要意义[8]; 其它方面, 盐角草对于滨海湿地重金属污染、有机物污染和海水富营养化的修复都有重要的作用[9]。
目前, 关于盐角草的研究主要集中在耐盐机理、形态结构、生理生化以及耐盐基因等方面, 对于盐角草种群相关的研究, 国内外还未见报道。因此, 本研究主要利用动态生命表以及相关性分析的方法来研究盐角草种群的动态变化及其与温度之间的关系, 以期为探析这一盐生植物在高盐干热极端环境条件下的耐盐机理提供依据。
1 材料与方法
1.1 研究区自然概况
研究区位于新疆维吾尔自治区吐鲁番市艾丁湖地区。艾丁湖是中国大陆的最低点, 海拔-154.31 m, 地理坐标为42° 32'-42° 43' N、89° 10'-89° 40' E。气候类型属于大陆性暖温带荒漠气候, 年均气温14.1 ℃, 极端高温达49.6 ℃, 年降水量16 mm, 蒸发量3 000 mm。独特的地形造成该地区独特的气候条件, 不仅干旱, 而且多大风, 多干热风天气[10]。
1.2 研究方法
1.2.1 样地的确定与调查 在2015年3月初, 于艾丁湖地区盐角草自然分布区内(42° 38'26″-42° 38'45″ N, 89° 15'52″-89° 15'07″ E)依据其往年的水分条件、周围环境、盐角草生长情况等因素确定出A、B、C三个典型样地:A样地地形偏高, 4月之后地表无积水, 盐角草密度相对较低; B样地位于地表径流覆盖范围内, 地表长期有积水, 盖度相对中等; C样地地表早期有积水, 种群密度相对最大, 盖度最高。样地确定好后, 在每个样地中分别设定3个0.5 m× 0.5 m样方进行调查, 根据盐角草的生长情况, 从3月至10月每月调查1~2次, 共10次, 每次统计样方内盐角草的株数。
1.2.2 动态生命表 生命表是研究种群随时间动态变化的有效方法[11]。一般有两种类型:一种是静态生命表, 一般指在某一特定时间内对种群作年龄结构的调查资料编制, 用空间代替时间的方法来分析种群年龄结构的动态变化; 另一种为动态生命表, 是根据同年出生的所有个体存活量动态监测资料编制而成[12]。其内容包括以下几个指标:在x时间段内出现的个体数(nx), 其值实际测得。在x时间段开始标准化存活的个体数(lx)(一般标准化为1 000), lx=nx/n0× 1 000。活到x天的比率(sx), sx=nx/n0。从x年龄级到x+1年龄级间隔期内标准化死亡数(dx), dx=lx-lx+1。从x年龄级到x+1年龄级间隔期内标准化死亡率(qx), qx=dx/lx。标准化存活数的自然对数值(lnlx)。各年龄组的致死力(kx), kx=lnlx-lnlx+1。
本研究参照醉香含笑(Michelia macclurei)[12]和梭梭(Haloxylon ammodendron)[13]当年生幼苗的动态生命表的编制方法, 自3月开始观测样方内盐角草存活个体数量, 每月统计一次(在结果期至枯黄期每月统计两次), 直至10月底全部死亡, 共统计10次。以每两次采样之间的时间段作为一个龄级, 共9个龄级。
1.2.3 存活曲线和死亡率曲线的绘制 存活曲线是依据存活个体数量来描述特定时期内的死亡率, 通过对特定时期个体数量相对作图得到。按Deevey(1947)[13]的划分, 一般有3种基本类型。Ⅰ 型是凸曲线, 属于该类型的种群绝大多数都是能活到该物种自然死亡年龄, 但当活到一定生理年龄时, 短期内几乎全部死亡; Ⅱ 型是直线, 也称对角线型, 属于该型的种群各年龄死亡率基本相同; Ⅲ 型是凹型曲线, 早期死亡率高, 一旦活到某一年龄, 死亡率就较低。本研究分别以3块样地各自的标准化存活量(lx)和标准化死亡率(qx)为纵坐标, 以龄级为横坐标绘制3个盐角草种群的存活曲线和死亡率曲线图。
1.2.4 温度的测量 鉴于每次采样时所测得的温度数据并不能代表当时一段时间内温度变化的实际情况, 因此温度数据以“ 中国天气网” 所公布的当地数据为主(http://www.weather.com.cn), 采样时所测得的温度作为参考, 来计算盐角草整个生命周期内温度变化的情况。本研究确定了4个温度指标— — 日最高温(观测日的最高温度)、日最低温(观测日的最低温度)、平均日最高温(以观测日前、后7天的日最高温数据算平均数)和平均日最低温(以观测日前、后7天的日最低温数据算平均数)。
1.3 数据统计分析方法
本研究利用Excel 2010进行数据处理和作图, 采用SPSS 19.0软件对各个被测指标值进行相关性分析。
2 结果与分析
2.1 种群变化特点
2.1.1 A种群动态生命表 A种群大小在3月至5月呈增长趋势, 标准化存活个体数由2 229上升到4 110, 增长率达到84%, 并在5月上旬达到峰值, 之后A种群便开始衰亡; 其中, 5月中下旬至6月初是衰亡的第1个高峰, 标准化存活个体数由4 110下降到2 610, 死亡率为37%, 致死力为0.45; 7月初至8月初是衰亡的第2个高峰, 也是衰亡最快的时期, 标准化存活数由2 356骤减为93, 死亡率高达96%, 几近100%死亡, 致死力达到3.23, 根据同时期的温度变化曲线, 可推测主要的致死因素可能是高温导致(表1)。A种群个体能存活到9月末的比率为0.025, 能存活到10月份的比率也只有0.025, 而盐角草一般9月中下旬开始开花, 10月份才开始结种, 因此, 存活率如此之低使得整个A种群种子库大小急剧缩减, 危及到该种群的可持续繁殖。另外, 除去10月份的正常死亡外, 其它整个时期A种群的总死亡率为97.5%, 而8月份之前的死亡率之和就达到90.7%, 因此, A种群衰亡过程主要发生在8月份之前, 8月份之后整个种群趋于稳定。
| 表1 盐角草A种群动态生命表 Table 1 Dynamic life table of Salicornia europaea population in plot A |
| 表2 盐角草B种群动态生命表 Table 2 Dynamic life table of Salicornia europaea population in plot B |
2.1.2 B种群动态生命表 B种群大小在3月末至4月末有个小滑坡, 之后又开始增长, 在6月初达到峰值(表2)。其中, 增长速度在4月至5月之间最大, 标准化存活比率由893增长到1 245, 增长率为39%。在6月峰值过后, B种群便开始衰亡, 其中, 7月至8月衰亡速度最快, 标准化存活个体数由1 207骤降为553, 死亡率为54%, 致死力为0.78。之后的衰亡趋势较为平缓, 死亡率保持在10%以下。除去10月份的正常死亡外, 整个B种群的总死亡率为45.5%, 8月份之前的死亡率之和为44.7%, 因此, B种群大小的急剧变化同样发生在8月份之前, 8月份之后整个种群的大小趋于稳定, 直到10月底全部自然死亡。
| 表3 盐角草C种群动态生命表 Table 3 Dynamic life table of Salicornia europaea population in plot C |
2.1.3 C种群动态生命表 C种群从3月末开始, 整个种群大小便持续衰减, 只在4月到5月之间有一个小回升, 增长率为12%, 之后便较为稳定, 平均死亡率保持在20%(表3)。除去10月份的正常死亡之外, C种群的总死亡率为79.6%, 较为均匀地分布于各时间段。因此, 相交于A、B种群来说, C种群更稳定, 更能适应周围环境的变化。
2.1.4 3个种群的存活曲线 整体来看, A种群的存活情况较为复杂, 5月上旬之前标准化存活个体数持续增长, 种群大小达到峰值, 之后的存活曲线属于Deevy-Ⅲ 型, 即刚开始时死亡率较大, 9月之后趋于稳定(图1)。B种群也属于Deevy-Ⅲ 型, 刚开始时死亡率较大, 8月之后趋于稳定。C种群属于Deevey-Ⅱ 型存活曲线, 呈对角线型, 即各时间段存活率基本一致。
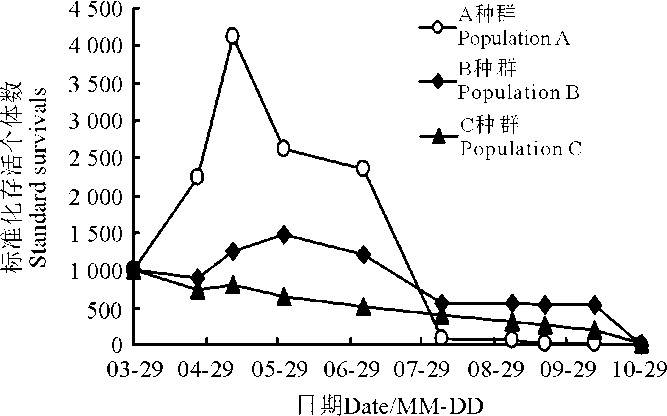 | 图1 3个盐角草种群的存活曲线Fig. 1 Survival curves of the 3 populations of Salicornia europaea |
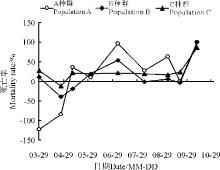
2.1.5 3个种群死亡率曲线 就死亡率而言, 除10月份的自然死亡外(图2), A种群有3个死亡率较高的时间, 第1个是在5月末至6月初; 第2个在7月至8月之间; 第3个在9月份。与A种群类似, B种群死亡率从5月末开始快速增大, 在5月末至6月初达到19%, 之后继续增大, 但增大的速度有所下降, 并在7月至8月间达到最大值54%; C种群只有1个死亡率较高的时期, 是在5月末至6月初, 之后死亡率基本一致, 一直保持在20%左右, 直到10月底全部自然死亡。
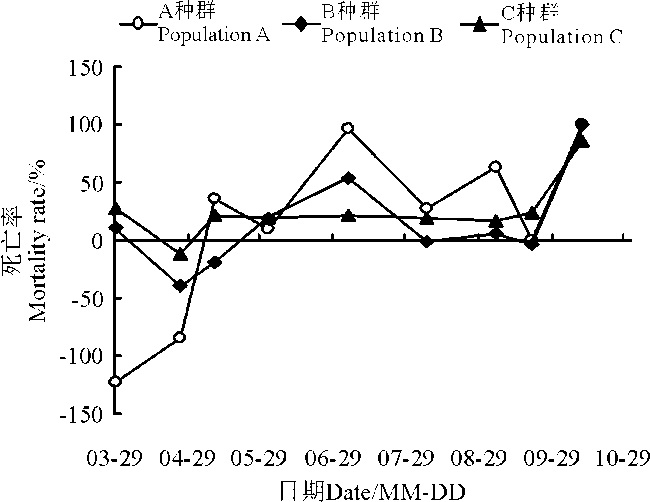 | 图2 3个盐角草种群的死亡率曲线Fig. 2 Mortality rate curves of the 3 populations ofSalicornia europaea |
2.2 温度变化特点
以观测日前、后7 d的数据计算日平均温度, 从3月29日到8月7日, 温度持续上升, 日最高温度从22 ℃上升到43 ℃, 平均日最高温由19.7 ℃上升到41 ℃; 日最低温度由9 ℃上升到26 ℃, 平均日最低温由11 ℃上升到26.3 ℃(图3)。之后便开始持续下降, 到10月31日时日最高温为17 ℃, 平均日最高温为19.1℃, 日最低温度为4 ℃, 平均日最低温为6.6 ℃。其中, 8月7日前后出现43 ℃左右的高温。
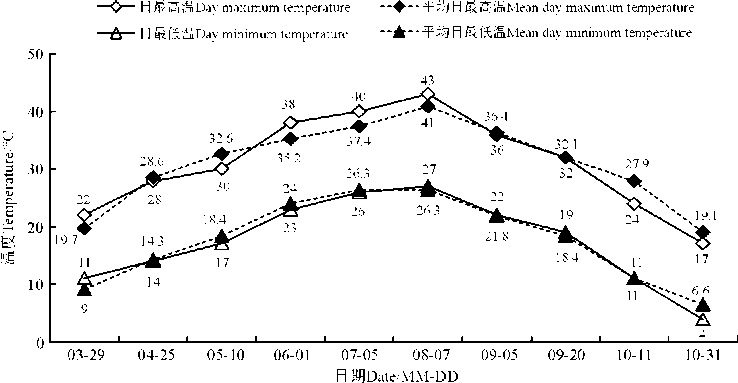 | 图3 研究期间的气温变化Fig. 3 Changes in the temperature during the study period |
2.3 温度变化与死亡率之间的相关性
将5月10日至9月6日的日最高温度与相应的盐角草死亡率做相关性分析(表4)可知, 在5月至9月期间, A种群死亡率与温度呈显著正相关关系(P< 0.05), 温度越高, 死亡率越大; B种群死亡率与温度呈极显著正相关关系(P< 0.01), 相关性系数达到0.990, 可见高温对B种群死亡率影响甚大; C种群死亡率与温度有一定相关性, 但不显著(P> 0.05)。
| 表4 温度与死亡率之间的相关性 Table 4 Correlation analysis between temperature and mortality rate |
3 讨论
在艾丁湖相同的大区域气候背景和不同的微域环境条件的共同作用下, 不同样地的盐角草种群的动态变化既有相似性, 也具有差异性。春季随着气温回升以及融雪对水分的补充, 水分及温度条件开始变得适宜于种子萌发。这一时期盐角草种子开始大量萌发, 种群密度快速增高并达到峰值。在5月前所有样地的盐角草种群密度均呈增长趋势。但在不同样地的小环境条件的影响下, 种群密度差异明显。A种群从3月至5月种群密度持续增长, 在5月10日左右达到峰值(1 940 plant· m-2)。于B种群而言, 从3月至4月种群密度持续增长, 达到3 720 plant· m-2, 随后出现自疏现象, 种群密度下降。而后随着温度的继续回升和积水的持续增加, 土壤养分条件更好, 种子继续萌发, 种群大小开始稳定增长, 到6月初种群密度达到峰值(4 424 plant· m-2)。C样地水分条件最好, 积水程度明显多于A样地, 种群密度持续增长, 5月达到了9 928 plant· m-2。3个种群相比, A种群密度低于B、C种群, 这可能与A样地较少的土壤积水有关。因为盐角草密度与土壤盐分负相关[3]。土壤积水越多, 一方面可以直接稀释盐分, 另一方面也可以缓解由于蒸发导致地下水上升引起的土壤返盐, 因此种群密度高, 这一结论也与该样地观测结果一致:A样地积水最少, 种群密度最小; C样地土壤积水最多, 种群密度最大。
之后, 在5月末至6月这段时期内, 最高温度从30 ℃升高到38 ℃以上, 而这一时期土壤水分和土壤营养物质含量呈现下降趋势(数据未发表), 3个样地的种群均进入第1次死亡高峰期。研究表明, 高气温可以使根际温度过高, 严重影响根系功能的发挥[14], 而此时38 ℃以上的温度有可能导致根温的上升, 从而影响盐角草的正常生长。因此, 这一时期, 温度的升高、水分下降、土壤营养物质减少可能形成了共同的胁迫作用, 从而引发了种群自疏, 导致了第1次的死亡率高峰期的出现。
在差异性方面, 7月至8月是A、B两个种群的第2个死亡率高峰期, 而C种群仍然维持着较低的死亡率。在这段时期内, 有连续长达34 d的40 ℃以上的极端高温天气。相较于C种群而言, A、B种群密度小, 致使土表裸露在外, 土壤温度对太阳辐射强度的变化敏感, 长时间的极端高温和较剧烈的昼夜温差不但影响盐角草根部对营养物质的吸收, 破坏盐角草体内正常的新陈代谢, 而且高温使得土壤水分蒸发强烈, 从而引起土壤返盐[15], 盐壳积聚于土表之上, 阻碍盐角草的生长, 最终导致A、B种群出现第2次死亡率高峰期。C样地土壤水分含量高, 环境胁迫程度低, 且C种群有较高的密度, 高密度可以使土壤电导率和无机盐含量显著下降[16], 并且使整个土表被盐角草覆盖起来, 这不但在一定程度上保持了土壤水分和土壤温度的恒定, 而且将土壤的水分蒸发由植物蒸腾取代, 从而减少土壤返盐, 降低耕作层中的盐分[15], 利于植物生长。另外, 高密度意味着土壤中的残枝败叶较多, 增加了土壤中的有机质, 有机质的增加使得土壤微生物增多, 从而改善了土壤的肥力和物理性状[17], 这反过来又提高了C种群的发芽率, 从而形成一个良性循环。因此, 这一阶段在相同的外界高温条件下, 温度对不同种群的影响不尽相同, 低种群密度使得高温的胁迫作用加剧, 而较高的土壤含水率以及较高的种群密度则大大缓解了高温的影响。
在9月, 温度下降到36 ℃左右, 高温胁迫程度有所缓和, 环境致死力下降。积水较多且种群密度较大的B、C种群可能已适应此时的环境条件, 因此死亡率保持平稳。而水分含量最少、种群密度最小的A样地可能依旧面临温度引起的土壤返盐形成的盐胁迫, 因此出现了第3次死亡率高峰期。
共同大环境的作用和小环境条件的差异性最终导致3个种群表现出不同的特征, A、B两个种群死亡率较高, 直至9月之后趋于稳定, 存活曲线属于Deevy-Ⅲ 型; C种群存活率高, 死亡率一直维持在较低水平, 在整个生命期内基本保持一致, 属于Deevey-Ⅱ 型存活曲线。
4 结论
艾丁湖地区盐角草种群对高温胁迫和盐胁迫的抵抗能力不只与个体的生物学特性有关, 还与环境以及种群特点有关。环境含水量高, 种群密度大地区的盐角草对高温胁迫和盐胁迫的抵抗能力强, 死亡率平缓, 存活曲线属于Deevey-Ⅱ 型; 环境含水量差, 种群密度小地区的盐角草对高温胁迫和盐胁迫的抵抗力弱, 在整个生命周期内有3个高死亡率时期, 存活曲线属于Deevy-Ⅲ 型。
研究表明, 影响植物物种空间格局的环境因素除气候因子之外, 还有地形要素和土壤性状[18], 因此, 土壤、地形等环境条件对盐角草种群动态的影响还有待于进一步研究。
(责任编辑 武艳培)
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|