{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
干旱胁迫对4种决明属植物光合作用和叶绿素荧光特性的影响
[姚春娟, 郭圣茂 , 马英超, 赖晓莲, 杨肖华]
, 马英超, 赖晓莲, 杨肖华]
, 马英超, 赖晓莲, 杨肖华]
|
|


第一作者:姚春娟(1990-),女,陕西宝鸡人,在读硕士生,研究方向为森林培育。E-mail:1105876426@qq.com
以4种决明属植物草决明( Cassia obtusifolia)、望江南( C. occidentalis)、伞房决明( C. corymbosa)、双荚决明( C. bicapsularis)为材料,采用盆栽控水试验,设置轻度干旱、重度干旱和正常灌水处理,研究了干旱胁迫对4种决明属植物的叶绿素含量、光合作用参数和叶绿素荧光特性的影响。结果显示,1)干旱胁迫导致4种决明属植物的叶绿素含量下降,望江南的下降幅度最小,说明望江南在水分减少的情况下仍具有较高的光合作用能力,伞房决明的叶绿素含量在3种处理下都趋于最高水平,表明伞房决明光能吸收、转换和传递能力较强;2)干旱胁迫导致4种决明属植物气孔导度(Gs)降低,气孔限制值(Ls)升高,伴随着胞间CO2浓度(Ci)降低,净光合速率(Pn)下降,表明是气孔因素造成的,此后,除草决明外其它3种植物的Ci均升高,Ls均下降,这是非气孔因素造成的;在干旱胁迫下望江南具有较高的光合速率,伞房决明具有相对较高的气孔开放度;3)干旱胁迫导致4种决明属植物的最大荧光(Fm)和光化学效率(Fv/Fm)下降,初始荧光(Fo)上升,表明PSⅡ结构受到破坏或者植物进行了热耗散。光化学淬灭系数(qP)和电子传递速率(ETR)下降,而非光化学淬灭系数(NPQ)上升,说明干旱胁迫对PSⅡ造成伤害,但其能够有效地避免或减轻因PSⅡ吸收过多光能而引起的光抑制和光氧化。望江南表现的最稳定,说明望江南具有较强的抗旱性。
The effects of drought stress on the characteristics of photosynthesis and chlorophyll fluorescence of four species of Cassia were studied by conducting a water control experiment (mild drought, severe drought, and normal irrigation) using four Cassia species ( C. obtusifolia, C. occidentalis, C. corymbosa, C. bicapsularis). The results showed that 1) drought stress resulted in a decline in the chlorophyll content in all four species of Cassia, and the stress degree of C. occidentalis was the lowest, meaning that C. occidentalis maintains the highest photosynthetic capacity under low moisture conditions. The chlorophyll content of C. corymbosa was the highest in all three treatments, indicating that the light energy absorption and transformation capacity of C. corymbosa are stronger than those of the other three species. 2) Drought stress resulted in all four species of Cassia displaying reduced stomatal conductance (Gs), while the stomatal limitation (Ls) value increased, which was accompanied by a decrease in intercellular CO2 concentration (Ci) and a decline in the rate of photosynthesis (Pn), indicating that drought stress affects stomatal factors. The exception to this was C. obtusifolia, which showed an increase in Ci and decrease in Ls, indicating that the results are because of non-stomatal factors. Under drought stress, C. occidentalis showed a higher photosynthetic rate, and C. corymbosa showed a relatively high degree of stomatal opening. 3) Drought stress led to a decrease in Fm and Fv/Fm, and an increase in Fo among the four species of Cassia, indicating that damage to photosystem Ⅱ(PSⅡ) or heat dissipation. The qP and ETR values decreased, while the NPQ value increased, indicating damage to PSⅡ by drought stress, but effectively avoiding or reducing photoinhibition and light oxidation, which is caused by PSⅡ absorbing excess light energy. C. occidentalis showed the most stable performance, which means that C. occidentalis has a higher drought resistance
干旱造成植物水分缺失, 是许多经济作物周期或长期性遭受的一种逆境胁迫[1, 2]。江西省地处长江中下游南岸, 雨水充沛, 但分布不均, 差异较大, 尤其表现在梅雨季节过后会出现持续干旱, 降水量稀少, 这对湿热敏感的决明属植物而言, 要想长期存活下去, 培育抗旱的决明属植物显得十分必要。决明属(Cassia)是豆科植物, 乔木、灌木或草本, 500~600种, 广泛分布于亚洲、非洲和美洲热带, 在我国南方和北方均有种植, 大约20余种[3]。该属大部分植物主要以种子入药, 如:草决明(C. obtusifolia)和双荚决明(C. bicapsularis)具有明目、治眼疾, 益肾、解蛇毒, 轻身、助肝气, 保健、去脂肪等功效[4-5]; 望江南(C. occidentalis)具有肃肺、清肝火, 止咳、消肿毒, 通腹、利便尿等功效。而伞房决明(C. corymbosa) 和双荚决明因花形美、颜色艳, 耐干旱、耐贫瘠, 易固氮、防虫伤等优点而广泛应用于园林景观中[6]。此外, 该属其它植物也发挥着重要作用。
光合作用是植物生长的基础, 而光合作用在很大程度上受到环境因素的限制, 其中, 干旱胁迫是植物常见的一种。叶绿素荧光作为一种植物内在的探针, 其参数变化能够有效反映外界环境对植物自身光合生理的影响。目前, 有关逆境胁迫下光合作用和叶绿素荧光特性的研究早已在水稻(Oryza sativa)、大豆(Glycine max)、玉米(Zea mays)等作物中广泛开展。研究表明, 光合作用和叶绿素荧光特性是作物抗逆性生理评价的重要指标, 作物在逆境胁迫下通过改变自身结构机制利用无机物生产有机物并且贮存能量, 这对掌握逆境中作物生理生长变化有指导意义, 而叶绿素荧光特性能客观表明叶片结构和组织之间的相关信息, 能及时反映胁迫条件下叶片响应的信息[7]。但近年来关于决明属植物的报道主要集中在临床应用、化学成分、药理作用等方面[4], 针对该属植物光合作用和叶绿素荧光特性的探究鲜有报道。为此, 对干旱胁迫下4种决明属植物光合作用和叶绿素荧光特性进行探讨, 阐明其与决明属植物之间的关系, 建立更准确、便捷地培育决明属植物耐旱性的方法和技术, 以期为决明属植物的抗旱育种提供理论参考。
选取前一年成熟的4种决明属种子(草决明、望江南、伞房决明、双荚决明)为供试种子, 于2015年3月开始进行盆栽育苗。试验方法具体如下:采用高22 cm、上口径为27 cm、下口径为17 cm的塑料盆进行盆栽试验, 各盆中分别装入7 kg园内风干黏壤土, 供试土壤含有机质1.62%, 全氮0.057%, 田间最大持水量为25.7%, pH 7.3。塑料盆底铺入等量鹅卵石, 鹅卵石上置滤纸, 达到与土隔离的目的, 盆中间插入硬质塑料管至鹅卵石上, 用于浇水。播种时对种子进行筛选, 选取籽粒健康饱满、无虫蛀鼠咬的种子, 浸泡于蒸馏水12 h, 捞出后稍晾干, 于2015年4月中旬播种, 每盆播5~6粒种子, 出苗后待苗长达到15 cm时每盆留苗3株。育苗期间进行正常的水分管理, 保证每周浇水2~3次。
试验在江西农业大学中药园内进行, 该地处于南昌市北郊, 28° 46'N, 115° 55'E, 亚热带大陆性气候, 4月-8月平均气温25.5~27.8 ℃, 4月-8月平均日照时数1 820~1 845 h, 自然环境优越。
2015年4月-8月在中药园防雨棚内对草决明、望江南、伞房决明、双荚决明幼苗进行盆栽控水试验, 通过控制水分达到干旱胁迫的目的, 其水分的控制以相对含水量为标准。待苗长达到40 cm时开始进行控水, 自然干旱进行至设定土壤含水量的标准范围内后, 每天08:00 和18:00 均采用称重法补水控水并且进行记录, 每个品种设计3个水平的处理。正常灌水(CK)相对含水量为田间最大持水量的75%~85%、轻度干旱(MS)为55%~65%、重度干旱(SS)为35%~45%, 各处理重复3次, 在处理15 d后, 选取受光相同、叶龄和叶位相对较一致的成熟型功能叶片作为被测对象, 每株选取测量3个叶片。
生长指标的测定:用电子数显卡尺测定植物基径, 用钢卷尺测定株高和冠幅。
叶绿素含量测定:采用日本柯尼卡美能达SPAD-502PLUS叶绿素仪进行测量, 测定时选取无叶脉的部位, 每片叶子至少选择15个点进行测量, 叶片面积较大的, 增加其SPAD的采集点, 求其平均值。
叶片相对含水量测定:将新采的植物叶片迅速剪成小块, 立刻称取两份1 g(鲜重)叶片, 一份放入80 ℃恒温烘箱中烘烤1 h, 称其干重; 另一份放入蒸馏水中浸泡70 min, 达到恒重时, 擦掉表面多余水分, 称其饱和重。计算公式为:
相对含水量=
光合作用气体交换参数测定:采用美国Li-COR公司生产的LI-6400型光合作用仪, 选择一个晴天上午, 于09:00-12:00进行测量。测定时选择Li-6400-02B红蓝光光源叶室, 温度为25 ℃, CO2浓度为400 μ mol· mol-1, 使用开放式气路, 空气的相对湿度为50%~70%, 设定有效光合辐射(PAR)为800 μ mol· (m2· s)-1, 测定的参数有净光合速率(Pn), 气孔导度(Gs), 胞间CO2浓度(Ci), 蒸腾速率(Tr), 计算气孔限制值(Ls=1-Ci/C0, C0为400 μ mol· mol-1)。
叶绿素荧光动力学参数测定:采用德国WALZ公司生产的便携式调制荧光仪PAM-2500, 选择09:00-12:00进行测量。测定前叶片进行暗适应20 min, 用笔做出标记保证每次能夹到相同位置, 测定的荧光参数主要有:初始荧光(Fo)、最大荧光(Fm)、光化学效率[Fv/Fm=(Fm-Fo)/Fm]、光化学淬灭系数(qP)、非光化学淬灭系数(NPQ)、表观光合电子传递速率(ETR)。
采用SPSS 17.0软件, 用单因素ANOVA(Duncan’ s新复极差法分析显著性, 用平均值± 标准差表示, 运用WPS Office MicrosoftExcel 2016制作图表。
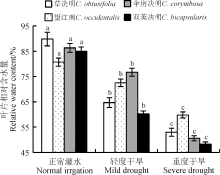
叶片相对含水量反映了植物体内水分亏缺的程度。随着干旱胁迫的增强, 4种决明属植物的叶片相对含水量均呈显著递减趋势(P< 0.05)(图1)。望江南较其它植物而言, 下降趋势较缓慢, 其降幅为25.9%, 处于最低水平。双荚决明在干旱胁迫下, 叶片相对含水量一直位于最低值, 其降幅最大, 为43.4%。草决明和伞房决明在正常灌水时叶片相对含水量较高, 受到轻度胁迫后, 草决明和伞房决明的降幅分别为28.1%和11.2%, 草决明降幅大于伞房决明, 而重度胁迫后, 降幅继续下降13.1%和30.2%, 伞房决明降幅大于草决明, 但总的降幅相差不大, 处于中间水平。
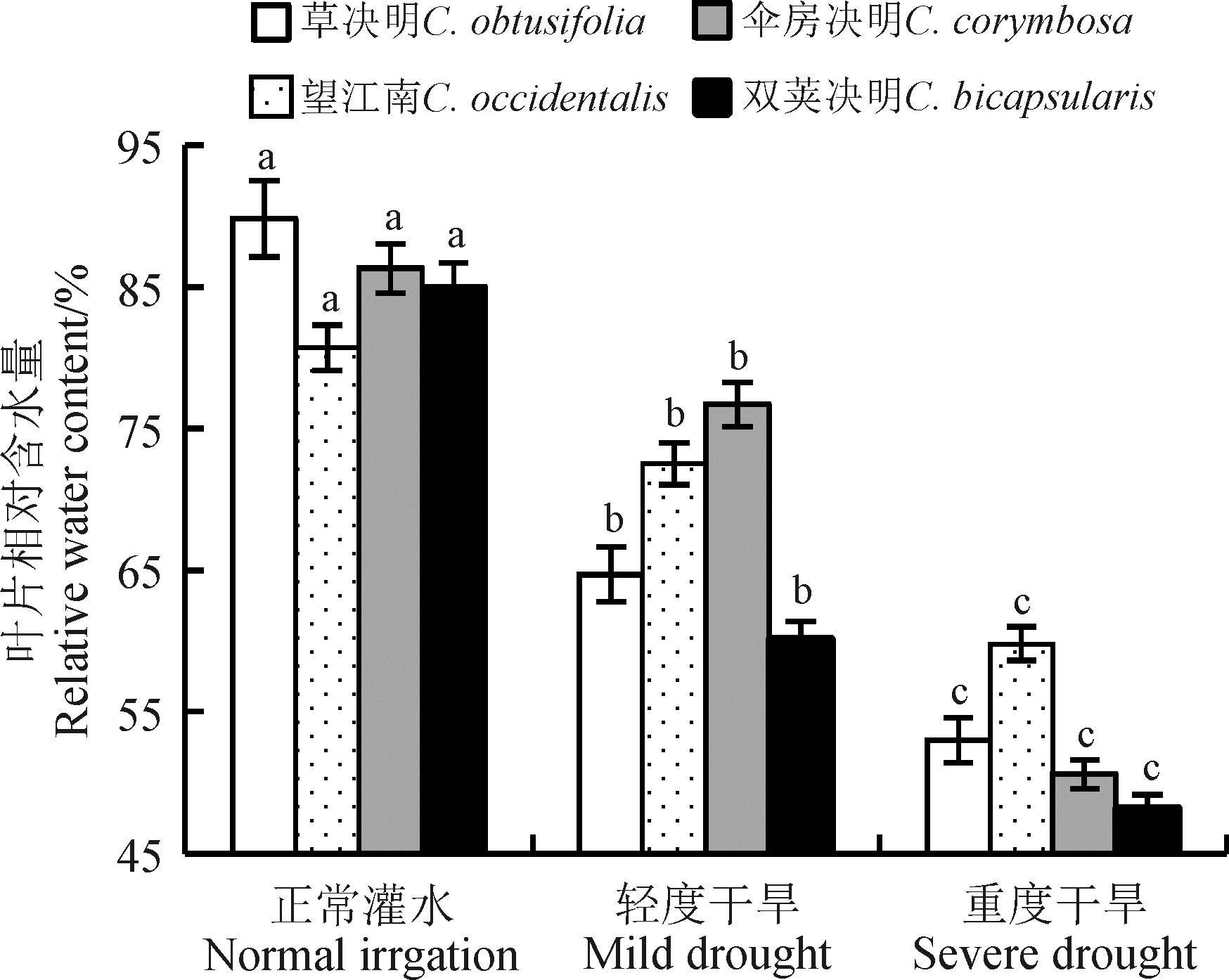 | 图1 干旱胁迫下4种决明属植物叶片相对含水量 注:不同小写字母表示不同干旱胁迫程度之间差异显著(P< 0.05)。下同。Fig. 1 Leaf relative water content of four species of Cassia under drought stress Note: Different lowercase letters indicate significant difference among different drought stress treatments at the 0.05 level. similarly for the following figures. |
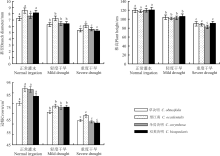
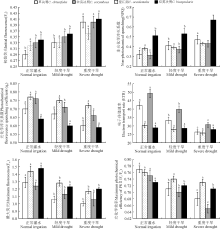
干旱胁迫下植物的基径、株高、冠幅是反映植物自然生长状况的3项重要指标。本研究中4种植物的基径、株高、冠幅在干旱胁迫下, 都呈递减趋势(图2), 随着干旱程度的加剧, 表现出显著差异(P< 0.05)。在干旱加剧的过程中, 望江南的基径、株高和冠幅的降幅最小, 分别为26.1%、25.6%和41.5%, 且各干旱程度时望江南的基径和冠幅为4种植物中的最高值。正常灌水下, 伞房决明的株高为最高值, 随着干旱胁迫的加剧, 在重度干旱下降低了31.1%, 冠幅降低58.4%, 在4种决明属植物中降幅最大。
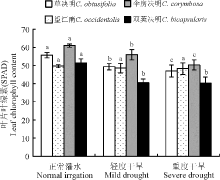
叶绿素是绿色植物进行光合作用的主要色素及重要物质基础, 其含量直接影响到植株的生长状况, 与叶片的光合能力密切相关[7, 8, 9]。SPAD值(叶色值, leaf colorvalues)能快速精确地表达植物叶片中叶绿素的相对含量[10, 11, 12]。干旱胁迫导致4种决明属植物叶片的SPAD值都有所下降(图3), 随着胁迫程度的加剧, 草决明和伞房决明均达到显著差异水平(P< 0.05), 而望江南的差异不明显(P> 0.05), 干旱胁迫处理的双荚决明显著低于正常灌水。根据SPAD值显示, 在不同干旱处理下, 伞房决明的SPAD值较其它3种植物而言处于最高水平, 而望江南的SPAD值降幅最小, 为2.82%。
 | 图2 干旱胁迫下4种决明属植物的生长特性Fig. 2 Growth characteristics of four species of Cassia under drought stress |
 | 图3 干旱胁迫下4种决明属植物叶片的SPAD值Fig. 3 Leaf SPAD value of four species of Cassia under drought stress |
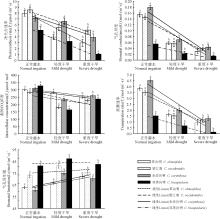
4种植物受干旱胁迫后, Pn、Gs、Tr均呈下降趋势(图4), 且随着干旱程度的加剧, 各指标均持续下降, 降幅排序分别为:伞房决明> 望江南> 双荚决明> 草决明; 双荚决明> 望江南> 伞房决明> 草决明; 双荚决明> 望江南> 伞房决明> 草决明。草决明的Gs、Tr、Ls, 望江南的Ci、Ls, 伞房决明的Gs、Ci、Tr, 双荚决明的Pn、Ci在不同干旱处理下均呈显著差异(P< 0.05)。望江南、伞房决明、双荚决明的Ci随着受胁迫程度的加重呈现出先降低后上升的趋势, 轻度胁迫下这3种植物的Ci处于最低水平; 4种植物Ci的降幅排序为草决明< 伞房决明< 望江南< 双荚决明; 在干旱程度加剧下, 4种决明属植物的Ls都有所上升, 且草决明的Ls一直上升, 而其它3种植物都呈现出先上升后下降的趋势。4种植物Ls升幅排序为双荚决明< 草决明< 望江南< 伞房决明。
2.5.1 最大光化学效率 干旱胁迫导致4种决明属植物的Fo升高, 且增幅随着胁迫程度的加剧而增大, 其中望江南的增幅最低, 为11.1%(图5)。干旱胁迫导致Fo增加, 表明PSⅡ 反应中心遭受了破坏或者可逆性的失活[13]。在干旱胁迫处理下, Fm均呈下降趋势, 望江南和伞房决明的降幅较小, 表明其在干旱胁迫下具有较高的PSⅡ 光化学活性。Fv/Fm也随着干旱程度的加剧呈下降趋势, 重度干旱与对照正常灌水相比, 4种决明属植物依次下降13.33%、3.95%、13.33%和2.74%。其中, 草决明和伞房决明的差异最大, 表明其叶绿素荧光受干旱胁迫影响较大, PSⅡ 反应中心受到损伤, 原初光能转化效率下降。
 | 图4 干旱胁迫对4种决明属植物的光合作用参数的影响Fig. 4 Photosynthesis parameters of four species of Cassia plant under drought stress |
 | 图5 干旱胁迫对4种决明属植物叶绿素荧光参数的影响Fig. 5 Chlorophyll fluorescence parameters of four species of Cassia under drought stress |
2.5.2 荧光猝灭动力学 4种决明属植物的qP在干旱胁迫下均有所下降, 且随着胁迫程度的逐渐增加, 望江南的降幅最小(图5)。qP下降说明降低了PSⅡ 反应中心的氧化态次级电子受体(QA)与开放部分的比例, 从而抑制QA向QB光合电子传递[14]。望江南在各处理中qP较高, 表明在干旱胁迫下望江南的QA重新氧化的能力较强, 其反应中心受体电子传递的破坏程度较弱。随着胁迫程度的加剧, 4种决明属植物的NPQ均呈上升趋势, 表明PSⅡ 反应中心耗散过剩光能的能力逐渐增强, 伞房决明的NPQ增幅高于其它3种, 表明其吸收的光能进行热耗散的较多, 而望江南的NPQ增幅较小, 说明其耗散较少, 光能的转化率较其它几种植物高。4种决明属植物随着干旱胁迫程度的增加, ETR均呈下降趋势, 但降幅不同。望江南和双荚决明受干旱胁迫的影响不大, 草决明和伞房决明的不同干旱处理之间均差异显著(P< 0.05), 受干旱影响较大。
干旱胁迫下, 植物叶片的相对含水量是植物保水能力的体现, 抗旱性强的植物有较强的保水能力。本研究中, 干旱胁迫下4种决明属植物的叶片相对含水量降低, 说明水分供应亏缺时, 植物用于自身构建和生理过程的水分也相应的减少, 水分的供求不平衡[15]。就不同植物而言, 干旱胁迫下望江南较其它3种植物的叶片相对含水量降幅最小; 就不同处理而言, 正常灌水下4种决明属植物叶片相对含水量排序为草决明> 伞房决明> 双荚决明> 望江南, 中度胁迫下为伞房决明> 望江南> 草决明> 双荚决明, 重度胁迫下为望江南> 草决明> 伞房决明> 双荚决明, 这表明望江南虽然自身叶片含水量较低, 但其水分调节能力较强, 较其它3种植物而言是一种具有较强保水能力的抗旱性植物。
从生长角度来看, 干旱胁迫下4种决明属植物的基径、株高、冠幅均呈显著递减趋势, 说明水分是限制植物生长的重要因素。本研究中, 同一干旱处理下, 基径和冠幅的最大值为望江南, 最小值为草决明, 株高的最大值为伞房决明; 随着干旱胁迫程度的加剧, 望江南的各生长指标降幅最小, 伞房决明的株高和冠幅降幅最大。表明望江南能较强的适应水分供应不足的环境, 在水分胁迫时能及时调节自身各器官之间的支配, 从而将较多的生物量分配到地上叶和枝的生长上, 从而有效地维持光合作用, 以满足自身在干旱环境中生长的需要[16]。
光合作用过程中叶绿素从光中吸收能量传递给反应中心, 所以叶绿素是参与植物光合作用最重要的因素, 与光合作用密不可分。当植物受到逆境胁迫时, 叶绿素含量的多少能在一定程度上反映植物的同化能力。本研究中, 干旱胁迫下4种决明属植物的叶绿素含量都有所降低, 说明干旱胁迫可导致叶绿素合成受阻或叶绿素降解。其原因可能是干旱胁迫下, 叶片细胞内水分减少, 叶绿素合成速率变慢会影响核糖体的形成, 蛋白质合成受阻, 代谢减慢, 抑制叶绿素的生物合成, 叶绿素被分解[17]。望江南虽然在正常条件下叶绿素含量最低, 但随着胁迫程度的加强, 到达重度胁迫后, 其叶绿素含量仅居第二, 而其叶绿素含量下降幅度最小, 这说明望江南在水分减少的情况下仍具有较高的光合作用能力, 能较强地维持叶片细胞体内光合碳素同化活动的正常进行, 叶片衰老较慢。另外, 伞房决明在干旱胁迫下的叶绿素含量高于其它3种植物, 表明伞房决明在光合作用的光反应阶段光能吸收、转换和传递能力较强, 比其它3种植物高。
光合作用是植物生长的生理基础, 能够准确反映植物遭遇水分亏缺时的生长势和抗逆性。植物在逆境胁迫下导致光合速率降低主要取决于气孔和非气孔因素, 它的判断标准主要是根据Ci和Ls的变化方向, 即Ci降低和Ls升高, 表明是气孔因素; Ci升高和Ls降低, 表明是非气孔因素[18]。本研究中, 4种决明属植物在干旱胁迫下, Gs降低, Ls升高, 伴随着Ci降低, Pn下降, 这表明是气孔因素造成的, 这是因为干旱胁迫使植物的叶片气孔收缩, 导致Gs降低, 使CO2由外向内扩散的阻力增加, 同时也限制了碳固定的底物的增加[19]。在此之后, 除草决明外其它3种植物的Ci升高, Ls下降, 引起这个现象的原因可能是非气孔因素, 随着干旱程度的逐渐加剧, 缺水导致叶片光合机构破坏或产生光抑制, 碳的同化能力降低。望江南在干旱胁迫的条件下, 具有较高的光合速率, 而在干旱加剧的情况下, 其Pn、Gs、Tr降幅最低, 说明望江南的光合器官受到的伤害较小, 光合细胞的活性较高。伞房决明在干旱胁迫下具有相对较高的气孔开放度, 其Pn、Gs、Tr、Ci的降幅均位于4种植物的从小到大排序的第二位。由此表明, 望江南和伞房决明都表现出良好的抗旱性。
叶绿素荧光作为植物光合作用的探针, 能够灵敏和准确地检测植物光合机构在环境胁迫下的响应。本研究中, 干旱胁迫导致4种决明属植物的Fm和Fv/Fm下降, 说明干旱造成植物的叶片发生了光抑制。与此同时, Fo上升, 表明PSⅡ 光合机构受到了损坏或者是植物本身一些防御性的激发能热耗散过程加强的反映[19, 20, 21]。望江南的Fo变化差异不明显, 说明干旱胁迫对望江南PSⅡ 反应中心造成的破坏较小, 其叶绿素吸收的光能还是以进行光化学反应为主。4种植物的Fm均降低, 胁迫与未胁迫对照之间均有差异, 说明干旱胁迫能够使PSⅡ 反应中心QA的氧化态数量减少。在干旱胁迫下4种植物的Fv/Fm也都呈现出降低的趋势, 说明干旱胁迫阻碍了植物光合电子传递, 导致自身化学能ATP和NADPH的合成受阻, 影响碳反应中的CO2同化。望江南的Fv/Fm在干旱胁迫下差异最不明显, 说明干旱胁迫下望江南能维持PSⅡ 较高的光合电子传递和光化学活性正常进行。其中草决明和伞房决明的差异明显, 表明其叶绿素荧光受干旱胁迫影响较大, PSⅡ 反应中心受到损伤, 原初光能转化效率下降。干旱胁迫导致4种植物的qP和ETR下降, 而NPQ上升, 说明干旱胁迫对PSⅡ 造成伤害, 但其能够有效地减免因PSⅡ 吸收太多光能而产生的光抑制和光氧化[18]。其中, 望江南的qP、NPQ、ETR表现的最稳定, 说明望江南够保持较高的光能利用效率和光合作用潜力, 热耗散较少, 碳同化能力和PSⅡ 激发能利用效率较高。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
干旱胁迫对4种决明属植物光合作用和叶绿素荧光特性的影响
姚春娟, 郭圣茂, 马英超, 赖晓莲, 杨肖华
Effect of drought stress on characteristics of photosynthesis and chlorophyll fluorescence of four species of Cassia
Yao Chun-juan, Guo Sheng-mao, Ma Ying-chao, Lai Xiao-lian, Yang Xiao-hua