

{kind=link}
{kind=link}
紫花苜蓿响应低温胁迫转录因子的鉴定及表达分析
引用本文
齐晓, 闵学阳, 张正社, 孙启忠, 刘文献. 紫花苜蓿响应低温胁迫转录因子的鉴定及表达分析. 草业科学, ,34(9):1824-1829
Qi Xiao, Min Xue-yang, Zhang Zheng-she, Sun Qi-zhong, Liu Wen-xian. Identification and expression analysis of transcription factors in alfalfa under low temperature stress. Pratacultural Science,,34(9): 1824-1829
Permissions
Qi Xiao, Min Xue-yang, Zhang Zheng-she, Sun Qi-zhong, Liu Wen-xian. Identification and expression analysis of transcription factors in alfalfa under low temperature stress. Pratacultural Science,,34(9): 1824-1829
Copyright©, 《草业科学》编辑部
《草业科学》编辑部
紫花苜蓿响应低温胁迫转录因子的鉴定及表达分析
第一作者:齐晓(1982-),男,河北泊头人,农艺师,在读博士生,主要从事草品种管理与推广工作。E-mail:tq07mms@sina.com
共同第一作者:闵学阳(1991-),男,甘肃张掖人,在读硕士生,主要从事牧草分子生物学研究。E-mail:minxy15@lzu.edu.cn
摘要
研究证明,转录因子广泛参与植物的生长发育过程并响应多种生物/非生物胁迫信号途径。低温胁迫可导致紫花苜蓿( Medicao sativa)减产、越冬率降低以及生产年限缩短。利用新一代高通量转录组测序技术,本研究对4 ℃低温胁迫下紫花苜蓿中响应低温胁迫的转录因子基因进行了鉴定,并对其表达情况进行分析,结果表明,转录组测序共获得78 925条Unigene序列和3 448个差异表达基因。其中,从3 448个差异表达基因中共鉴定出43个转录因子家族,共251个基因被显著地诱导表达。不同转录因子家族基因受低温胁迫诱导表达不同。本研究有助于在整体水平加深了解紫花苜蓿转录因子表达特性,为进一步研究这些响应低温胁迫的转录因子的功能提供参考。
关键词:
紫花苜蓿; 转录因子; 低温胁迫; Unigene; 转录组
中图分类号:S816
文献标志码:A
文章编号:1001-0629(2017)09-1824-06
doi: 10.11829/j.issn.1001-0629.2016-0552
Identification and expression analysis of transcription factors in alfalfa under low temperature stress
Abstract
Numerous studies have shown that transcription factors are important in regulating plant development and responses to various biotic/abiotic stressors. Low temperature stress could result in lowered production, reduced overwintering rate and a decrease in sustainability. In the present study, the identification and gene expression model of the transcription factors in alfalfa under low temperature stress (4 ℃) were analysed by the high-through put RNA-Seq technology. The results showed that a total of 78 925 Unigene sequences and 3 448 differently expressed genes were produced in our study. There were 43 transcription factor families, and a total of 251 genes were identified from all differently expressed genes. The expression model of these differently expressed transcription factor genes was different. This study facilitates the understanding of the expression model and verifies the related function of these low temperature response transcription factor genes in alfalfa.
Keyword:
Medicago sativa; transcription factors; low temperature stress; Unigene; transcriptome
由于植物的相对固定和难移动性, 在面对诸多生物和非生物逆境胁迫(干旱、低温、病虫害等)时, 无法像动物一样通过移动来规避不利环境的影响。因此, 在长期的演化过程中, 植物进化出了复杂多样的防御体系来应对极端环境的侵害[1, 2]。在这些防御体系中, 多种转录因子家族发挥着关键调控作用, 日益受到人们的关注[3, 4, 5]。
转录因子是一类能够与真核生物基因顺式作用原件相互作用从而激活或抑制下游被调控基因转录的DNA 结合蛋白质[6]。截至目前, 已有至少64个转录因子家族得以鉴定和研究[7]。越来越多的研究也证明, 转录因子通过自身以及与其它蛋白质互作, 进一步调控下游基因的转录表达, 从而广泛参与响应植物干旱、高盐、低温、激素、病虫害以及调控生长发育等生命过程[8, 9]。因此, 深入研究和鉴定参与响应生物/非生物逆境胁迫转录因子的功能及其调控途径, 对提高植物抗逆性以及抗性育种具有重要的指导意义。
紫花苜蓿(Medicao sativa)是全球种植面积最大的豆科牧草, 具有广泛的生态适应性、稳定的生产力以及较高的营养价值, 在我国农牧业生产和生态经济建设中发挥着巨大作用[10, 11]。但研究发现, 低温胁迫可导致紫花苜蓿减产、越冬率降低以及生产年限缩短, 极大地限制了紫花苜蓿在高寒地区的种植和推广[12, 13]。模式植物研究结果证明, 在深入解析植物响应低温胁迫分子调控机制的基础上, 利用基因工程技术以及常规抗寒品种的选育是提高植物响应低温胁迫的有效途径[14]。但截至目前, 紫花苜蓿响应低温胁迫的分子机制仍不清楚, 迫切需要对其内在分子调控机制进行深入研究。因此, 本研究利用转录组高通量测序和生物信息学技术, 对紫花苜蓿中响应低温胁迫的不同转录因子家族进行系统筛选和分析, 以期为揭示紫花苜蓿响应低温逆境胁迫的分子机理以及苜蓿抗寒品种选育提供理论依据。
1 材料和方法
1.1 试验材料和处理条件
试验材料采用美国紫花苜蓿冬季存活能力(winter survival)鉴定标准品种ZG9830(抗寒等级为一级, 最抗寒), 种子由美国农业部国家种质资源库(U.S. National Plant Germplasm System)提供。将种子置于营养土(土壤∶ 草炭∶ 蛭石=3∶ 2∶ 1)中育苗(育苗盆直径8 cm、深10 cm, 育苗30盆, 每盆3~5株), 在位于农业部全国草业产品质量监督检验测试中心的人工气候室进行(育苗条件为20 ℃恒温, 16 h/8 h, 昼/夜)。待幼苗长至3-5叶期(约40 d龄)时, 转至德国RUMED-3401型低温光照培养箱进行低温胁迫(4 ℃, 16 h/8 h, 昼/夜, 降温速率为每分钟1 ℃), 获取低温处理48 h的叶片样品。以 20 ℃条件下生长的叶片样品作为对照(CK)。降温前, 幼苗在低温培养箱中(20 ℃恒温, 16 h/8 h, 昼/夜)适应48 h。取下的叶片经液氮处理后保存于- 80 ℃备用。各处理设3次生物学重复, 每次生物学重复由3~5株幼苗的8~10片三出复叶(不包括最顶端新叶和有明显非低温损伤的叶片)组成。取样时均处于光照条件下。
1.2 RNA提取及转录组测序
分别利用TRIzol(Invitrogen, 美国)和RNeasy Mini试剂盒(Invitrogen, 美国)对样品总RNA进行提取和纯化。分别采用Nanodrop、Qubit 2.0、Aglient 2100方法, 检测RNA样品的纯度、浓度和完整性等, 以保证使用合格的样品进行转录组测序。6个样品获取到4.6~17.9 μ g的RNA进行建库, 质量均满足建库要求, 总量均满足3次及以上常规量建库。北京百迈客生物科技有限公司利用Illumina HiSeq 4000高通量测序平台进行转录组测序, 测序读长为PE150。
1.3 测序数据的过滤、组装和注释分析
利用FASTX软件(http://hannonlab.cshl.edu/fastx_toolkit/)去除Raw Data中的接头和低质量序列后得到Clean Data, 并利用Trinity程序对进行组装最终获得Unigene序列。
利用BLAST程序(E-value=10-5)将Unigene序列分别与NR、Swiss-Prot、GO、COG、KOG以及KEGG数据进行比对, 获得Unigene的注释信息。
1.4 差异表达基因(differentially expressed genes, DEG)的筛选
采用Bowtie软件[15]将各样品测序得到的reads与Unigene库进行比对, 根据比对结果, 结合RSEM方法[16]进行表达量评估, 并利用每千碱基外显子百万片段数(fragments per kilobase of transcript per million mapped reads, FPKM)方法[17]计算样本间的基因表达差异。将低温处理较之未处理样品间表达量符合错误发现率(false discovery rate, FDR)≤ 0.01且|log2Ratio|≥ 1条件的基因定义为差异表达基因。
1.5 转录因子的鉴定
大豆(Glycine max)、蒺藜苜蓿(Medicago truncatula)及百脉根(Lotus japonicus)转录因子家族蛋白质序列下载于豆科转录因子数据库(legume transcription factor database, Legume TFDB)[18]。将3种豆科植物中的相同转录因子家族序列合并在一起, 利用BLAST程序(E-value=10-10)与Unigene进行比对, 鉴定紫花苜蓿中潜在的转录因子基因。
2 结果与分析
2.1 转录组测序与组装
对紫花苜蓿ZG9830低温胁迫处理0和48 h的叶片转录组测序后共获得78 925条Unigene序列。所有Unigene序列的N50值为1 329, 表明本研究的转录组组装完整性较高。Unigene的序列长度主要分布在300~3 000 nt之间, 共有76 164条Unigene序列, 占总序列的96.50%。Unigene序列长度大于3 000nt的序列有2 761条, 占总序列的3.50%。由图1可以看出, 本研究所获得Unigene序列数量随着测序长度的增加呈逐渐减少的趋势, 表明测试样品具有较好的测序质量。
 | 图1 Unigene序列长度分布Fig. 1 Length distribution of Unigene sequences |
2.2 Unigene功能注释
通过BLAST(E-value=10-5)比对分析, 78 925条Unigene共有43 930个Unigene在6个公共数据库中得到注释, 占总Unigene数的55.66%(表1)。其中, NR数据库中注释到的Unigene数目最多, 达到41 223个, 占93.84%; 最少的为KEGG数据库, 注释到的Unigene数目为12 760, 占29.05%。
| 表1 Unigene功能注释 Table 1 The functional annotation of Unigene |
2.3 差异表达基因的鉴定
以FDR≤ 0.01和|log2Ratio|≥ 1作为DEG筛选条件, 相比未低温处理的对照样本, 低温处理48 h后共得到3 448个DEG, 占总Unigene的7.85%。其中, 1 835个Unigene上调表达, 占53.22%; 1 613个Unigene下调表达, 占46.78%。
2.4 转录因子家族的鉴定
将所有紫花苜蓿叶片Unigene序列与大豆、蒺藜苜蓿和百脉根转录因子蛋白序列进行同源比对, 得到59个潜在的紫花苜蓿转录因子家族共1 730个基因。其中, 预测得到的转录因子基因数目最多的10个家族分别为PHD、ABI3VP1、AP2_EREBP、bHLH、atypical_MYB、HB、(R1)R2R3_Myb、WRKY_Zn、bZIP和NAC(表2)。紫花苜蓿经低温处理后, 3 448个DEG中共有43个转录因子家族共251个基因被显著地诱导表达。其中, 差异表达显著且包含转录因子数量最多的10个家族分别为PHD、AP2_EREBP、bHLH、atypical_MYB、WRKY_Zn、Myb_related、GRAS、NAC、HB和bZIP(表2)。
| 表2 紫花苜蓿全部及差异表达转录因子的数目 Table 2 Number of total unigenes and differentially expressed transcription factors |
2.5 响应低温胁迫差异表达转录因子表达分析
低温胁迫诱导下转录因子数量最多的10个家族的DEG的表达模式各不相同(图2)。上调表达的转录因子共有84个, 下调表达的转录因子共有60个。 其中, 转录因子WRKY_Zn家族(11个基因)和NAC家族(10个基因)都为上调表达。
3 讨论与结论
低温胁迫通过抑制植物生长发育从而制约植物的生态分布并直接影响农作物的产量和品质, 甚至造成绝收[19, 20]。因此, 对植物特别是重要农作物耐寒性及其机理进行研究具有重要的理论和实际应用价值。近年来随着研究的深入, 发现一些小RNA分子, 例如microRNA以及siRNA分子也参与到植物对低温胁迫的响应过程中[14, 21, 22]。另外, 转录因子在调控植物基因转录表达以及应答多种逆境胁迫过程中也发挥着重要的作用。可见, 植物对低温胁迫的响应是一个复杂的基因表达过程, 涉及到多种基因和信号转导途径[23]。
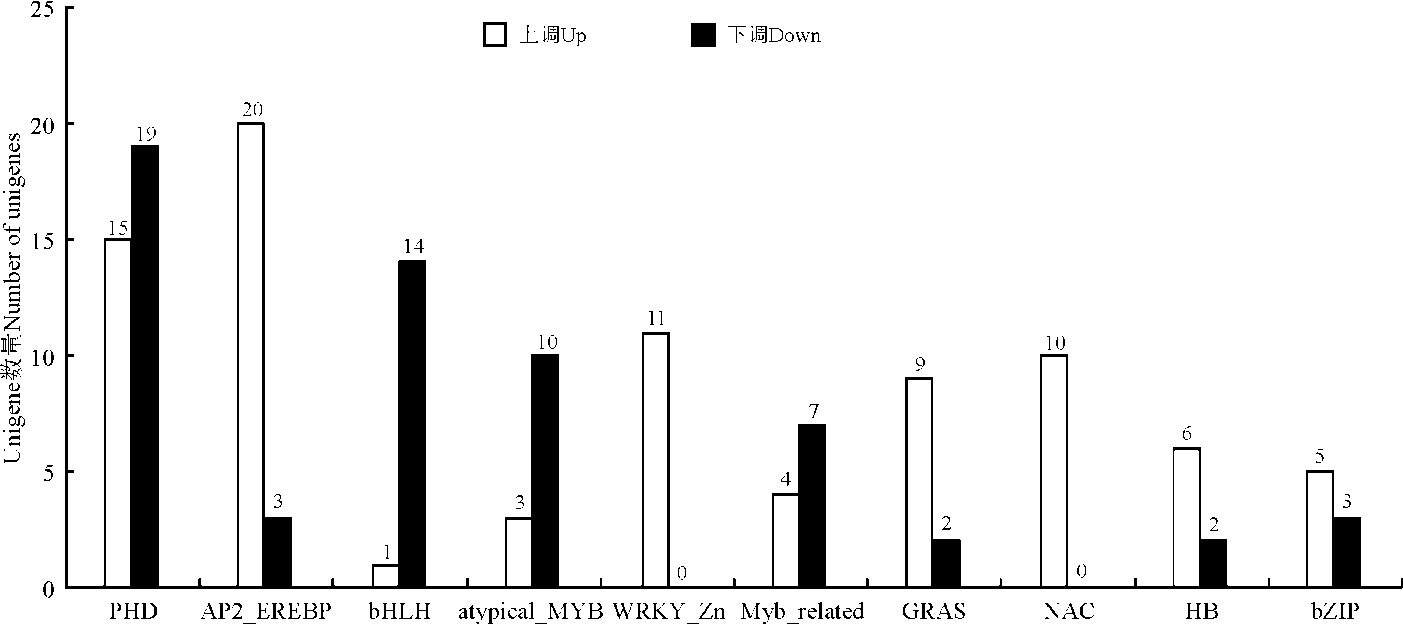 | 图2 紫花苜蓿响应低温胁迫转录因子表达Fig. 2 Expression of alfalfa transcription factors following low temperature stress |
随着后基因组时代的到来, 高通量测序技术已被广泛用于研究模式和非模式植物的生长发育以及对逆境胁迫响应的机制研究中[24, 25]。利用转录组测序技术, 陈嘉贝等[26]在盐胁迫下的两个甜瓜(Cucumis melo)品种中鉴定出19个和20个转录因子家族, 预测这些受盐胁迫调控的转录因子基因可能参与到甜瓜响应盐胁迫反应途径中。本研究通过对一个具有高抗寒级别的紫花苜蓿品种ZG9830进行4 ℃低温胁迫处理48 h后进行转录组分析, 共得到了78 925条Unigene序列和3 448个DEG。所有的Unigene序列中, 有93.84%的序列得到了注释(表1)。其余未被注释的序列可能是紫花苜蓿所特有的基因序列, 仍有待于进一步的分析。本研究中相关大量数据的获得有助于增加对苜蓿抗寒性机理的认识, 并可为后续抗寒相关基因的克隆及功能验证奠定基础。
通过BLAST比对分析, 从3 448个DEG中共鉴定出43个转录因子家族共251个基因被显著地诱导表达(表2)。截至目前, 已有多种转录因子家族, 包括MYB、AP2_EREBP、NAC、bHLH等参与调控植物响应低温胁迫反应[23]。例如, 在拟南芥(Arabidopsis thalianna)中过表达水稻OsMYB3R-2基因后, 转基因拟南芥植株的抗寒性显著提高[27]。NAC是一类植物特有的转录因子。水稻OsNAC6转录因子受冷处理诱导表达, 说明该转录因子参与水稻低温诱导信号转导途径[28]。水稻转录因子OsbHLH1被证明参与了冷胁迫的基因应答调控[29]。在本研究中, 差异表达显著且包含转录因子数量最多的10个家族分别为PHD、AP2_EREBP、bHLH、atypical_MYB、WRKY_Zn、Myb_related、GRAS、NAC、HB和bZIP。其中, bHLH转录因子家族15个基因中14个下调表达, 1个上调表达。atypical_MYB转录因子家族中3个上调表达, 10个下调表, 而NAC转录因子家族中的10个基因全部受低温诱导上调表达(图2)。可见, 这些转录因子也都参与了紫花苜蓿响应低温胁迫的调控途径; 而其它具有差异表达的转录因子家族如何参与调控紫花苜蓿低温胁迫还有待于进一步研究。
通过转录组测序及生物信息学分析, 在紫花苜蓿中获得了大量响应低温胁迫的差异表达基因, 并在转录组水平鉴定和分析了参与调控紫花苜蓿低温诱导反应的转录因子家族基因及其表达模式, 从而有利于从整体水平加深了解紫花苜蓿转录因子表达特性, 为进一步研究这些响应低温胁迫的转录因子的功能提供参考, 也可为紫花苜蓿耐低温功能研究提供新的基因资源。
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|