

{kind=link}
{kind=link}
{kind=link}
鸭茅EST-SSR标记在4种禾草上的可转移性
[王新宇, 张新全 , 黄琳凯, 聂刚]
, 黄琳凯, 聂刚]
, 黄琳凯, 聂刚]
|
|


第一作者:王新宇(1991-),男,四川成都人,在读硕士生,主要从事草种质资源创新及育种。E-mail:wxysicauDg@163.com
为了检验鸭茅EST-SSR标记在几种禾本科植物上的可转移性并筛选出可应用于后续研究的引物,本研究用40对可在鸭茅( Dactylis glomerata)上扩增产物的鸭茅EST-SSR引物,分析了6份多花黑麦草( Lolium multiflorum)、6份高羊茅( Festuca arundinacea)、3份扁穗牛鞭草( Hemarthria compressa)和3份牛鞭草( H. altissima)共18份材料的转移率、遗传多样性和亲缘关系。结果表明,多年生黑麦草和高羊茅可转移率为60%,扁穗牛鞭草和牛鞭草转移率最低,均为50%,26对引物共获得条带128条,多态性条带121条,多态性比(PPB)为94.53%,平均扩增出多态性条带4.92条,平均多态性信息量0.952。18份材料遗传相似系数范围为0.208~0.974,在遗传系数为0.68时,4个物种分别独自聚为一大类。本研究证实利用鸭茅EST序列开发的EST-SSR在有一定亲缘关系的物种间转移是可行的,可转移的引物对未来禾本科遗传育种研究有一定的参考价值。
In order to test the transferability of orchardgrass EST-SSR in several grass species in Gramineae and select some reliable markers used in followed-up studies, 40 orchardgrass EST-SSR, which can amplify products successfully in orchardgrass, were analysed to determine the transferability, genetic diversity and relationship of 18 materials including 6 Lolium multiflorum, 6 Festuca arundinacea, 3 Hemarthria compressa and H. altissima. The cross-transferability of these markers was found to be 60% in L. multiflorum and F. arundinacea, while 50% in H. compressa and H. altissima. Together, the 25 EST-SSRs detected 128 bands. In these 128 bands, 121 were polymorphic (94.53%) and the average number was 4.92 per primer, and the polymorphic information content (PIC) ranged from 0.75 to 1 with an average of 0.952. The similarity coefficient of the 18 materials ranged from 0.208 to 0.974, with an average of 0.490, and UPGMA cluster analysis grouped them into four groups at the similarity level of 0.68. This study confirmed that it was feasible that the EST-SSRs developed from orchardgrass EST sequences can cross to some related species. Those transferable markers will give some reference to breeding work in Gramineae.
SSRs即简单重复序列(simple sequence repeat), 又称微卫星DNA, 是一种由1~6个碱基为重复单位的DNA序列[1], 不论真核生物还是原核生物都有广泛且随机的分布。SSR标记与其它分子标记相比有明显的优势:SSRs有较高的丰度、较高的信息含量、较高的多态性、较多的共显性遗传和较多的重复性[2]。所以, 其已被广泛用于SSR遗传连锁图谱构建[3]、 DNA指纹图谱的构建[4]、遗传多样性的研究[5]、标记辅助选择育种[6]和品种鉴定[7]等。EST(Expressed Sequence Tag)-SSRs即表达序列标签是基于EST序列开发的新型分子标记, EST处在表达区域, 可以通过对比EST序列信息和蛋白质序列数据库推断基因功能[8, 9]。相比SSRs, EST-SSRs更保守, 所以在相关物种上EST-SSRs有更强的转移性[10]。设计多花黑麦草(Lolium multiflorum)、高羊茅(Festuca arundinacea)、扁穗牛鞭草(Hemarthria compressa)和牛鞭草(H. altissima)等非模式植物的SSR引物成本相对高一些, 但也可以增加这4个物种的标记种类。SSR能在物种之间转移是因为其侧翼序列的保守性, 但是SSR一般来自于非编码区, 物种之间SSR的差异大, 而EST序列来自于编码区, EST-SSR的侧翼序列高度保守, 所以EST-SSR能在物种之间有较强的转移能力。基于这个优势, EST-SSR在基因组学上有大量的应用。Wang等[11]用苜蓿(Medicago sativa)和大豆(Clycine max)的28个EST-SSR将百合属(Lilium)4个种26个品种成功聚为4类。忻雅等[12]研究白菜(Brassica pekinensis)EST-SSR对油菜(B. campestris)的转移性时发现, 28对引物有24对能在油菜上扩增出产物, 其中又有18对显示多态性。禾本科作物间转移性也有比较深入的应用:Tang等[13]研究小麦(Triticum aestivum)EST-SSR引物在大麦(Hordeum vulgare)、水稻(Oryza sotiva)和玉米(Zea mays)上的转移性发现, 转移性随亲缘关系变远而降低; Saha等[14]研究高羊茅EST-SSR在其它几种草上的可转移性, 并将同一引物扩增的产物进行测序, 发现扩增片段有很高的相似性; Xie等[15]用小麦EST-SSR和鸭茅(Dactylis glomerata)SSR研究了74份来自世界各地鸭茅的多样性和遗传关系, 结果表明鸭茅遗传多样性丰富。罗小燕等[16]使用柱花草(Stylosanthes)EST-SSR标记在山蚂蝗(Desmodium racemosum)上研究其遗传多样性, 发现两者有比较高的亲缘性, 柱花草EST-SSR可以使用在山蚂蝗的后续研究中。随着大量测序结果在网上公开, 开发EST-SSR能力得到极大地提升, 大量的植物可以利用已知EST序列信息开发引物, 并在自身和有亲缘性的物种上运用。但鸭茅EST的开发和运用目前还未见报道, 本研究利用鸭茅EST-SSR标记对黑麦草、高羊茅、扁穗牛鞭草和牛鞭草的转移性进行分析, 并尝试应用于这4种禾草亲缘关系和遗传多样性分析中, 为4种禾本科植物与鸭茅比较基因组学提供分子生物学依据, 并发掘优质基因, 为遗传育种提供帮助, 同时丰富这4个物种的分子标记数量。
本研究共用18份材料, 其中多年生黑麦草6份(编号1-6), 高羊茅6份(编号10-15), 扁穗牛鞭草(编号7-9)和牛鞭草(编号16-18)各3份, 但部分来自美国基因库的PI种质无法获得来源地信息, 具体情况如表1所示。供试材料叶片均采于四川农业大学草业科学研究基地, 采集后干燥保存。
每份种质随机选取健康幼嫩叶片, 采用试剂盒法(天根生化科技有限公司)提取DNA。最后用1%的琼脂糖凝胶电泳对提取的DNA浓度与纯度进行检测。符合实验要求的DNA样品放置于-20 ℃冰箱中保存备用。
引物由Huang等开发[17], 再经南京金斯瑞生物科技有限公司合成, 详细信息如表2所示。引物首先在‘ 金牛’ 、 ‘ 阿索斯’ 、 ‘ 01472’ 、 ‘ Intensiv’ 4个形态差异明显的鸭茅上进行筛选, 共选取40对扩增条带清晰、有一定多态性的引物用于鸭茅亲源物种上转移性分析。PCR 扩增反应总体积为15 μ L, 包括PCR-Mix(北京天根生化公司)7.5 μ L、模板DNA等1.5 μ L(10 ng· μ L-1)、正反向引物各0.6 μ L(10 pmol· μ L-1)、Taq酶0.3 μ L (2.5 U· μ L-1), ddH2O补足15 μ L。PCR反应程序:94 ℃预变性4 min, 94 ℃变性1 min, 退火温度1 min, 72 ℃延伸1 min, 共35个循环, 72 ℃下10 min, 4 ℃保存。扩增产物在6%聚丙烯酰胺凝胶上进行检测。样孔中每个样品点6 μ L, 以博瑞克公司的D50Marker为对照。330 V下电泳160 min。再用0.1%的AgNO3进行银染色和在NaOH液中显色, 凝胶在灯光下用数码相机照相保存以供分析。
| 表1 供试的禾草 Table 1 Four grass plant species examined in this study |
根据电泳结果, 在同一扩增位置, 有条带记为1, 无条带记为0, 建立0、1矩阵。数据由Office Excel标准化统计, 利用NTSYS-PC2.1中similarity程序计算相似系数, 以clustering程序中SHAN 进行UPGMA (un-weighted pair-group method with arithmetic means, 非加权组平均法)聚类分析。利用Popgen32计算多态信息含量(PIC)、Nei’ s遗传多样性指数(He)及Shannon 信息指数(I)。
本试验中40对鸭茅EST-SSR引物中在所有材料中扩增(图1, 图2)。其中有13对未扩增出条带(DGSSR 23、DGSSR 33、DGSSR 35、DGSSR 53、DGSSR 63、DGSSR 85、DGSSR 105、DGSSR 107、DGSSR 114、DGSSR 122、DGSSR 128、DGSSR 139、 DGSSR148); 另外27对引物可以在这4个物种上扩增出产物, 转移率达到67.5%。有17对引物在试验的每个的物种上均扩增出条带, 可转移率达到42.5%, 占可转移引物的63%。其中多花黑麦草可转移率为60%(24/40), 高羊茅可转移率为60%(24/40), 扁穗牛鞭草和牛鞭草转移率最低, 均为50%(20/40)。
| 表2 本研究中采用的鸭茅EST-SSR引物信息 Table 2 Dactylis glomerata EST-SSR primers used in the present study |
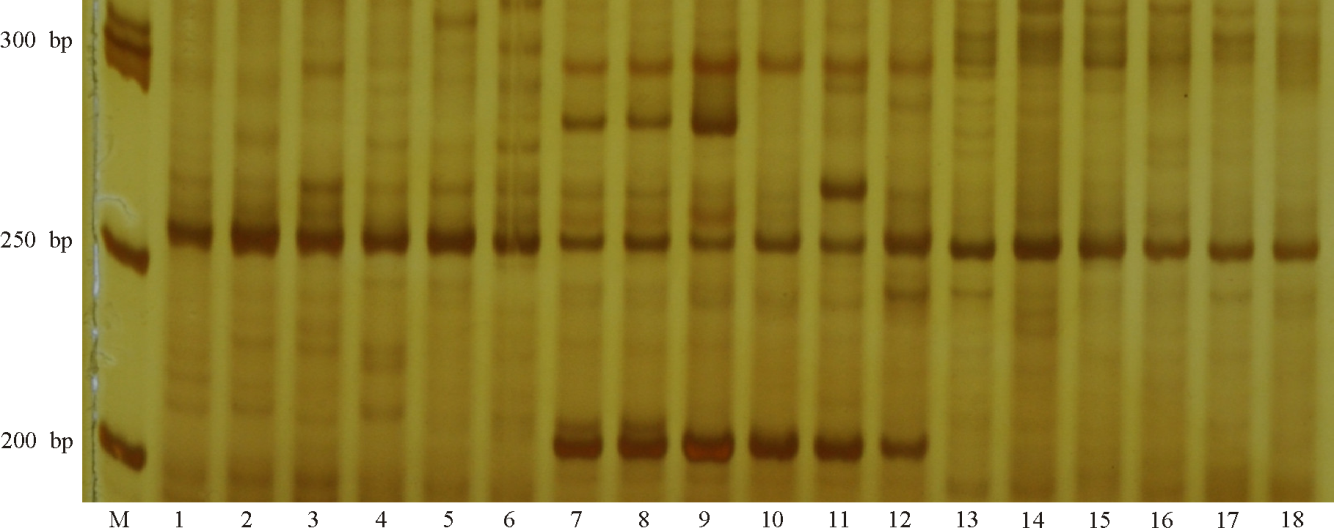 | 图1 引物DGSSR66在18份禾草扩增情况 注:M, marker; 1, 长江2号; 2, 阿德纳; 3, 达伯瑞; 4, 特高; 5, 安格斯1号; 6, 邦德; 7, PI549115; 8, PI434051; 9, PI561430; 10, PI578712; 11, PI574521; 12, PI520749; 13, 广益; 14, 重高; 15, 雅安; 16, H2O2; 17, H203; 18, H2O4。 下图同。Fig. 1 Amplified patterns of 18 grass species with primer pair DGSSR66 Note: M, marker; 1, Changjiang No.2; 2, Aderenalin; 3, Double Barrel; 4, Tetragold; 5, Angus 1; 6, Abundant; 7, PI549115; 8, PI434051; 9, PI561430; 10, PI578712; 11, PI574521; 12, PI520749; 13, Guangyi; 14, Chonggao; 15, Yaan; 16, H202; 17, H2O3; 18, H204. similarly for the following figures. |
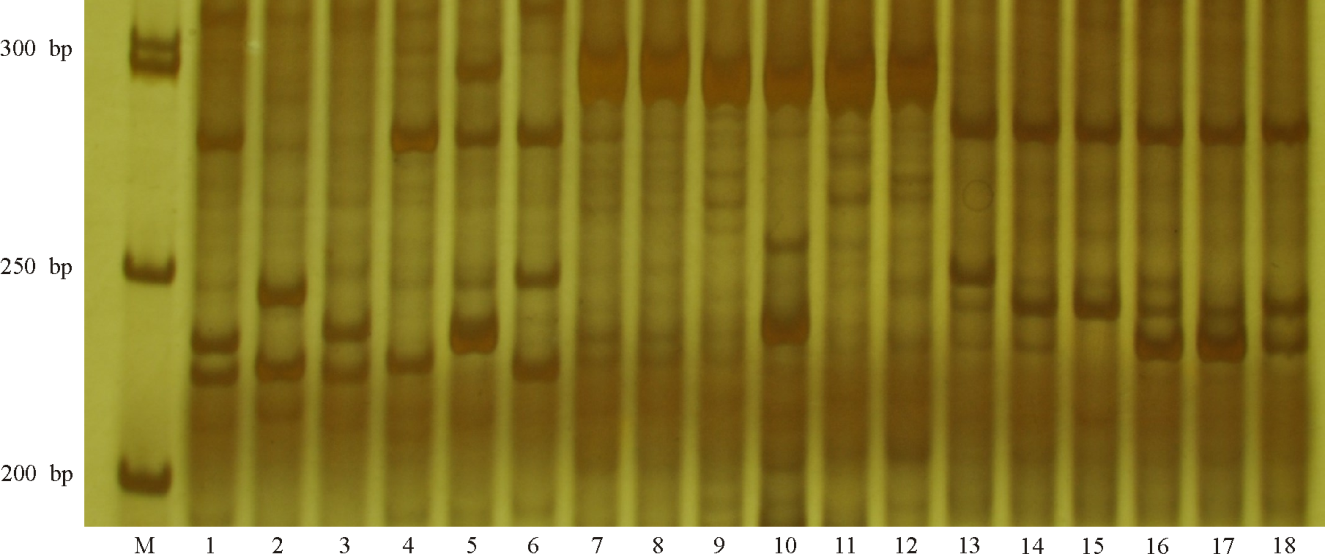 | 图2 引物DGSSR140在18份禾草扩增情况Fig. 2 Amplified patterns of 18 grass species with primer pair DGSSR140 |
26对(其中DGSSR82只在牛鞭草上出现扩增情况, 且无多态性)可转移的鸭茅EST-SSR引物共在4个物种上扩增出128条条带, 其中多态性条带为121条, 多态性条带比率为94.53%, 每对引物扩增出条带数为3~9条, 平均为4.92, 平均多态性条带数为4.65(表3)。每条引物多态性信息含量为0.75~1, 平均为0.951 6。4个物种的18份材料平均Nei’ s遗传多样性指数为0.306, 而平均Shannon信息指数为0.462。就各位点而言, Nei’ s遗传多样性指数最大为0.390, 最小为0.141, Shannon 信息指数最大为0.577, 最小为0.228。
| 表3 鸭茅EST-SSR标记在4种禾草中的多态性 Table 3 Polymorphism of Dactylis glomerata EST-SSR markers in four grass species |
运用NTSYS-PC2.1对数据进行遗传相似性分析, 结果表明, 4个物种18份材料遗传系数范围为0.208~0.974, 平均0.490(图3)。其中, 遗传相似系数最大的是雅安和广益(同为扁穗牛鞭草), 为0.974; 其次是PI520749和PI578712(同为高羊茅), 为0.937。这说明它们之间遗传关系很近。遗传相似系数最小的是安格斯1号(多花黑麦草)和H203(牛鞭草), 为0.191。4个物种18份材料遗传差异明显, 遗传多样性丰富。
基于遗传相似系数, 对4个物种18份材料进行聚类分析, 在遗传系数为0.680处可以将4个物种分成4类, 第Ⅰ 类为多花黑麦草, 第Ⅱ 类为高羊茅, 第Ⅲ 类扁穗牛鞭草, 第Ⅳ 类为高牛鞭草。从聚类分析图可以看出, 6个高羊茅品种的遗传相似系数较大, 亲缘关系较大; 其次是3个扁穗牛鞭草品种, 再次是6个多年生黑麦草品种, 最后是3个高牛鞭草品种。
 | 图3 4种禾草EST-SSR聚类分析图Fig. 3 UPGMA dendrogram of the four grasses species based on Dice’ s similarity coefficient according to orchardgrass EST-SSR markers |
EST来自表达序列, 与非编码区域相比, 它们在近缘物种上有更强的保守性, 所以EST-SSR比genomic-SSR存在更大的跨物种潜力[18]。而国内外大量的试验也证明, EST-SSR在物种间比较作图和比较基因组学上更有优势[19, 20]。近年来, 随着测序技术的发展以及成本的降低, 可以轻易地在各大数据库上得到大量的EST序列信息, 基于EST序列信息开发引物变得更加高效, 而EST-SSR引物在近缘种属间的转移性优势可以大大降低开发自身引物的成本, 弥补分子标记的不足, 丰富标记数量。本研究所用引物在4个物种上最低的转移率都达到50%, 证明利用鸭茅EST开发的引物运用在有一定亲缘性物种上的可行性, 本研究所用引物可以为以后与鸭茅有一定亲缘性的物种遗传多样性分析、品种鉴定、分子标记辅助育种、构建遗传图谱等提供新引物, 也为鸭茅与这4个有亲缘性的物种提供比较基因组学信息。
引物的转移性高低与两个物种间的亲缘关系有关。简而言之, 物种之间亲缘性越近, 引物转移性就越高; 反之, 亲缘关系越远, 引物转移性就越低[21]。Fan等[22]用新开发的梨(Pyrus)SSR标记在部分蔷薇科物种上研究转移性, 发现在同属苹果亚科的苹果(Malus pumila)的转移性达到58.2%, 在同属苹果亚科的枇杷(Eriobotrya japonica)的转移性达到32.8%, 而在与它亲缘关系较远的桃子(Amygdalus persica)上的转移性为14.1%, 在樱桃(Cerasus pseudocerasus)上为15.5%, 而在杏(Armeniaca vulgaris)上为12.7%。Varshney等[23]研究发现, 165条大麦EST-SSR引物, 78.2%可在小麦中转移, 75.2%可在黑麦(Secale cereale)中转移, 而只有42.4%可在水稻中转移, 转移性随着其种系关系距离增加而降低。鸭茅和高羊茅同属早熟禾亚科(Pooideae), 亲缘关系在4个物种中最近, 但在本研究中, 引物在多花黑麦草和高羊茅上的转移率均为60%, 试验结果不完全符合4个物种与鸭茅的亲缘关系情况。虽然转移率有差异, 但实际上可转移的引物数量差并不大。EST-SSR是否扩增产物也与DNA有关, DNA存在大量内含子或者SSR一端或者两端引物位于内含子、外显子剪切位点都会造成无效扩增, 估计高羊茅的的扩增率低于多花黑麦草是由以上两个原因造成的, 还有本研究所用引物数不多, 若引物达到一定数量, 高羊茅的转移率会有高于多花黑麦草的可能。
研究中18份材料可以被成功聚为4类, 证明转移性良好的引物对物种进化分析和物种鉴定都有重要意义。但是试验结果与黄婷等[24]和黄秀等[25]在多年生黑麦草、扁穗牛鞭草和高牛鞭草的聚类上和遗传相似性上均存在部分差异。黄婷等[24]的试验表明阿德纳和邦德的亲缘关系最近, 而本研究表明二者的亲缘关系很远。这可能是因为多年生黑麦草的遗传背景比较复杂, 而两次试验时间有近两年的距离, 材料会存在一定的差异, 且黄婷等使用30株单株DNA和30株混合DNA作为试验材料, 这两种混合方式产生的条带数会不同于单株材料的试验结果; 并且黄婷等[24]统计时条带也不是全部统计, 而是统计高于某一频率的条带, 这会忽略部分频率不是太高的条带; 两个试验的引物也不相同, 这些试验材料、统计方法和引物上的不同会直接导致聚类差异。黄秀等[24]的试验表明, 聚类结果完全和材料的生长地有关, 但本研究表明, 聚类结果不完全和材料的生长地有关。扁穗牛鞭草和高牛鞭草广泛地使用无性繁殖, 可以稳定地保持遗传特性, 试验材料没有太多的差异。两个试验都是EST-SSR引物, 但是黄秀等[25]使用的是禾本科通用引物, 而本研究使用的引物是基于鸭茅抗旱基因开发出来的, 这种具有一定功能的基因在有一定亲缘性地物种之间肯定会更加保守, 保守性会使条带多态性下降, 这在一定程度上也影响了聚类结果。本研究选用的引物数量较少, 不能完全说明4个物种的进化关系。本研究中多态性条带比率高达94.53%, 远高于板栗(Castanea mollissima)EST-SSR在栲树(Castanopsis fargesii)上的分析结果(56.6%)[26]和小麦在7种能源植物的分析结果(76.7%)[27], 说明本研究开发的引物可以利用在与鸭茅近缘物种上的亲缘性分析。
到目前为止, CNBI共收录EST序列信息7 600万条, 植物大概占1/3, 其中主要集中在水稻、小麦、玉米、大豆等作物, 而鸭茅等草类植物的EST信息相对较少。本研究结果进一步证明, EST序列有较高的保守性, 根据这一特性, EST信息可以更多的利用在重要基因定位以及重要基因开发利用上, 这对我国牧草分子育种大有裨益, 黑麦草、高羊茅、扁穗牛鞭草和牛鞭草属于优质草种, 在畜牧和草坪建植上都有广泛的使用, 经济价值也比较高[28, 29]。因此, 可以加大在鸭茅等草类植物EST相关研究。
利用40对鸭茅EST-SSR引物对本科4个物种18份材料进行转移性分析发现, 鸭茅EST-SSR引物的转移性随着与试验材料之间的亲缘关系变远而降低。本研究开发的鸭茅EST-SSR引物可以将18份材料按物种分为4类, 且扩增产物多态性高, 可用于鸭茅与有一定亲缘性的物种之间的后续研究。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
鸭茅EST-SSR标记在4种禾草上的可转移性
王新宇, 张新全, 黄琳凯, 聂刚
Transferability of Dactylis glomerata EST-SSR markers to four grass species
Wang Xin-yu, Zhang Xin-quan, Huang Lin-kai, Nie Gang