





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
放牧和AM真菌对夏季青藏高原东缘老芒麦生长和白粉病的影响
引用本文
李芳, 张峰, 郭艳娥, 高萍, 段廷玉. 放牧和AM真菌对夏季青藏高原东缘老芒麦生长和白粉病的影响. 草业科学, 2017,34(12):2528-2537
Li Fang, Zhang Feng, Guo Yan-e, Gao Ping, Duan Ting-yu. Influence of grazing and arbuscular mycorrhiza fungal infections on vegetative growth and powdery mildew incidence in Elymus sibiricus grown in the eastern margin of the Tibetan Plateau in summer . Pratacultural Science,2017,34(12): 2528-2537
Permissions
Li Fang, Zhang Feng, Guo Yan-e, Gao Ping, Duan Ting-yu. Influence of grazing and arbuscular mycorrhiza fungal infections on vegetative growth and powdery mildew incidence in Elymus sibiricus grown in the eastern margin of the Tibetan Plateau in summer . Pratacultural Science,2017,34(12): 2528-2537
Copyright©2017, 《草业科学》编辑部
放牧和AM真菌对夏季青藏高原东缘老芒麦生长和白粉病的影响
第一作者:李芳(1990-),女,青海西宁人,在读博士生,研究方向为牧草病理学。E-mail:lif2013@lzu.edu.cn
摘要
自玛曲3个放牧强度放牧地取土样,并采集优势植物老芒麦( Elymus sibiricus)种子,在温室条件下研究了3个放牧强度下土壤丛枝菌根(arbuscular mycorrhiza)真菌对老芒麦生长和白粉病的影响。放牧强度分别为轻度放牧、中度放牧和重度放牧,土壤处理分别为未作任何处理(自然土)、湿热灭菌土、湿热灭菌后还原除AM真菌外微生物土以及湿热灭菌后接种根内球囊霉( Rhizophagus intraradices)和幼套球囊霉( Claroideoglomus etunicatum)混合菌剂土。结果表明,放牧和AM真菌均可影响老芒麦生长和白粉病发生。老芒麦白粉病的发病率因不同放牧强度和土壤处理而异,中度放牧和重度放牧土壤处理下,老芒麦白粉病发病率低于轻度放牧处理,老芒麦总生物量、总磷含量、光合速率均高于轻度放牧土壤处理。老芒麦在自然土处理下白粉病发病率最高,为59.83%,灭菌土处理下老芒麦白粉病发病率最低,为44.73%。与自然土处理相比,接种AM真菌处理老芒麦白粉病发病率降低13.15%,总生物量增加56.46%。放牧强度和土壤处理对老芒麦总磷含量、叶绿素含量、多酚氧化酶存在交互效应。
关键词:
放牧强度; AM真菌; 逆境胁迫; 老芒麦; 白粉病
中图分类号:S812.8
文献标志码:A
文章编号:1001-0629(2017)12-2528-10
doi: 10.11829/j.issn.1001-0629.2016-0618
Influence of grazing and arbuscular mycorrhiza fungal infections on vegetative growth and powdery mildew incidence in Elymus sibiricus grown in the eastern margin of the Tibetan Plateau in summer
Abstract
We established a greenhouse experiment to study the influence of arbuscular mycorrhiza (AM) fungal infections on plant growth and powdery mildew incidence in the grass species Elymus sibiricus. Soil was collected from Gansu Maqu alpine grassland and used as follows: no treatment, sterilized to eliminate all microorganisms, sterilized and microorganisms returned except for AM fungi, sterilized and inoculated with a mix of Rhizophagus intraradices and Claroideoglomus etunicatum. E. sibiricus was grown on each of the treated soils and subjected to light grazing (LG), moderate grazing (MG), or heavy grazing (HG). Our analyses showed that growth and powdery mildew incidence in E. sibiricus was influenced by grazing and AM fungal colonization. Powdery mildew incidence varied among grazing intensities and soil treatments. Compared with the intensity in LG, in MG and HG, E. sibiricus had a lower powdery mildew incidence and higher total dry weight, total P content, and photosynthetic rate. The rate of powdery mildew was the highest (59.83%) in plants on soil without any treatment and the lowest (44.73%) in plants on sterilized soil. The rate of E. sibiricus powdery mildew disease decreased 13.15% and total plant dry weight increased by 56.46% after inoculation with AM fungi compared with no treatment. Grazing intensities and soil treatments had interaction effects on total P content, chlorophyll content, and PPO activity.
Keyword:
grazing intensity; arbuscular mycorrhizal fungi; stress; powdery mildew; Elymus sibiricus
真菌、细菌和放线菌等微生物大量存在于土壤中, 其中真菌种群生物量最高, 而丛枝菌根真菌(arbuscular mycorrhiza fungi, AMF)是土壤中生物量最大的一类微生物, 其广泛存在于草地、农田、森林等生态系统中, 可与约90%的维管植物根系形成共生体[1], 是世界上分布最广泛的一类菌根类型。我国AM真菌资源丰富, 截至2011年在我国已发现、报道的丛枝菌根真菌有10属131种, 占全世界已报道AM真菌25属250种的52.5%[2]。AM真菌能改善土壤理化性状[3]、改善根围微生物区系[4]、改变植物形态结构[5, 6]、促进植物水分和养分吸收利用[7, 8], 从而促进植物生长, 增加植物的产量。
放牧是天然草地主要的管理和利用方式, 也是影响草地植被的重要生态因子之一, 其对草地植物影响是多方面的[9, 10]。家畜通过釆食牧草、践踏和粪便3种主要途径改变草地植物种类组成和生物量等植被特征[11, 12, 13], 进而影响草地生态系统的群落结构和功能。此外, 在放牧条件下, 家畜的采食、践踏和排泄等行为对AM真菌也能产生影响, 进而影响菌根对牧草养分的吸收和生长发育的效应, 最终对草地农业生态系统植物群落特征和土壤微生物菌群产生深远的影响[14]。尤其是在过度放牧的地区这种影响尤为突出。研究者[15]发现放牧强度能影响AM真菌泡囊结构在大针茅(Stipa grandis)根系中的发育, 轻度和中度放牧可促进AM真菌泡囊的形成, 而重度放牧则显著抑制。对内蒙古草原不同放牧强度下AM真菌的变化进行研究发现, 随着放牧强度的增加, AM真菌的种类和孢子密度显著降低[16]。这可能是因为过度放牧时家畜的采食和践踏造成地表植物物种种类减少和盖度下降(物种种类减少了15种, 盖度下降60%), 从而导致植物向地下部输送的碳源不能满足大量AM真菌的生长需求[17]。但是放牧对AM真菌的影响因AM真菌的种类不同而存在差异, 如根内球囊霉(Rhizophagus intraradices)、地表球囊霉(Glomus versiforme)等对长期过度放牧反应敏感, 但另外一些种, 如摩西球囊霉(G. mosseae), 则不受长期过度放牧的影响[16]。
我国草地植物种类丰富, 同时草地病害发病也较严重。目前已知病害有2 831种, 其中, 禾本科牧草病害为1 289种, 占总数的45.5%, 是草本植物中病害最多的一科[18]。据报道, 早熟禾(Poa annua)白粉菌(Blumeria graminis)、长芒草(Stipa bungeana)锈病(Uredo sp.)[19]以及柱花草(Stylosanthes sp.)炭疽病菌(Colletotrichum gloeosporiodes)[20]和多年生黑麦草(Lolium perenne)感染冠锈病菌(Puccinia coronata)[21]均能造成牧草产量或种子产量降低, 甚至造成寄主植物大面积死亡[20]。
高山草原占我国草原总面积的三分之一, 甘肃甘南玛曲草原是典型的高山草原, 气候环境条件严酷, 草原在当地生态环境、经济建设、社会稳定和民族团结中具有重要的作用。老芒麦(Elymus sibiricus)分布在我国西北、华北、东北及青藏高原等地区, 具有适应范围广、抗旱抗寒性强、经济价值高的特点[22], 白粉病菌为专性寄生菌, 种类多, 分布广, 其引发的病害广泛存在于各种作物、蔬菜、花卉、树木等植物上。由于白粉病菌引起的病害大多为多循环病害, 病害潜育期短, 一个生长季节可繁殖多代, 且繁殖率高, 产生的孢子可借助空气大范围传播, 并多次重复侵染寄主植物, 所以一旦遇到合适的气候和环境条件, 此类病害就有可能大范围发生且流行快。寄主植物被白粉菌侵染后, 光合能力降低, 蒸腾强度增加, 严重阻碍植物的正常生长发育, 造成巨大的经济损失[23]。
作为青藏高原高山草原主要建群种之一, 老芒麦(Elymus sibiricus)根系能形成丛植菌根, 但共生体对植物养分吸收、植物生长及对普遍发生的白粉病的影响尚不清楚。本研究以禾本科牧草老芒麦为试验材料, 探究甘肃甘南玛曲草原3个放牧强度下土壤AM真菌对老芒麦生长过程中白粉病的影响, 以期通过放牧和利用AM真菌, 促进植物生长, 防控老芒麦白粉病, 从而提高草地牧草产量和品质。
1 材料和方法
1.1 试验地概况
试验采样地位于青藏高原东缘的甘肃省甘南玛曲草原站, 海拔为3 500 m左右。该区域属于高寒湿润气候, 年均温为1.2 ℃左右, 月均最低(1月) 和最高(7月)平均气温分别为-10 ℃和11.7 ℃, 年降水量约为620 mm。主体土壤类型为亚高山草甸土, 植被类型是禾本科的垂穗披碱草(Elymus nutans)、老芒麦、早熟禾、匍匐翦股颖(Agrostis stolonifera)、羊茅(Festuca ovina); 莎草科的线叶嵩草(Kobresia capillifolia)为优势种。小花草玉梅(Anemone rivularis)、珠芽蓼(Polygonum viviparum)、草原老鹳草(Geranium pratense)、莓叶委陵菜(Potentilla fragarioides)、丝叶毛茛(Ranunculus tanguticus)等为常见种。同时伴有其他杂草的典型高寒草甸草原。试验站优势植物种群除莎草科线叶嵩草不能和AM真菌形成共生体外, 其他建群种禾本科植物均可以和AM真菌形成共生体。
1.2 试验材料
1.2.1 供试土壤 将从玛曲放牧地所采集回来的土壤, 挑拣其他杂物并混合均匀后过筛, 筛子孔径为0.25 mm。土壤速效磷含量为5.16 mg· kg-1。
1.2.2 供试植物 供试植物为老芒麦(Elymus sibiricus), 种子从玛曲放牧地所采集。挑选大小一致且籽粒饱满的种子, 先用75%的酒精消毒3 min, 再用3%的次氯酸钠对种子进行表面消毒10 min, 用自来水和蒸馏水冲洗3次后, 均匀地摆放在装有湿润、无菌滤纸的培养皿内, 每皿25粒, 培养皿直径为9 cm, 置于黑暗条件下培养24 h后, 移至25 ℃光照培养箱中催芽48 h。
1.2.3 供试AM菌种 供试AM菌剂为根内球囊霉(Rhizophagus intraradices)和幼套球囊霉(Claroideoglomus etunicatum)混合菌剂, 购买于北京市农林科学研究院植物营养与资源研究所, 中国丛枝菌根真菌菌种资源保藏中心, 经三叶草(Trifolium pratense)扩繁所得。
1.3 试验设计
1.3.1 放牧地设置 玛曲放牧地分为轻度放牧(LG)、中度放牧(MG)和重度放牧(HG)3个放牧强度。轻度放牧为每轮牧小区8只羊, 共6个轮牧小区, 周期45 d; 中度放牧为每轮牧小区16只羊, 共6个轮牧小区, 周期45 d; 重度放牧为每轮牧小区8只羊, 放牧期间持续放牧, 不轮牧。暖季6-9月为放牧季节, 放牧期7-14 d(根据气候和牧草生长阶段适度调整)。每个轮牧小区选取若干植被盖度和群落相对一致的地点, 按照对角线取样法, 挖取5-15 cm土层土壤装入自封袋, 每个点3 kg左右。每个放牧强度采集约50 kg左右土样, 带回兰州进行温室试验。
1.3.2 丛枝菌根真菌处理设置 温室试验于2014年7月到2014年10月在兰州大学草地农业科技学院榆中智能温室中进行, 对各放牧处理过筛后的土壤进行以下4种处理:
自然土(NS):试验地土壤不经过任何处理, 保留原有的AM真菌及其他微生物区系;
灭菌土(SS):采用高温湿热灭菌法, 于121 ℃高压灭菌两次, 每次灭菌1 h, 两次灭菌间隔24 h;
灭菌后还原除AM真菌外微生物土(NM):土壤灭菌后(方法同上), 还原上述灭菌土壤中除AM真菌以外的其他微生物。将100 g自然土用200 mL无菌水稀释, 用两层42号滤纸过滤两次, 过滤掉AM真菌孢子, 滤液中除AM真菌以外的其他微生物, 将该滤液按10 mL· kg-1的比例加入无菌土, 即可得到不含AM真菌的土壤。
接种AM真菌土(AM):土壤灭菌后(方法同上), 装入灭菌土至花盆3/5处, 各称取15 g扩繁的幼套球囊霉和根内球囊霉混合菌剂平铺在上面, 然后再覆盖一层无菌土至4/5处。
1.3.3 试验管理 将以上处理好的土壤, 按照每盆700 g, 装入花盆, 按土壤干重的10%浇水。在装有上述各处理土壤的花盆中, 移栽催芽的老芒麦幼苗, 每盆移栽5株, 出苗一周后, 选择生长较为一致的保留3株, 生长期间根据水分损失情况, 补充至土壤重量的10%。生长期间每周测定一次株高和分蘖。植物在生长7周后, 自然发生白粉病, 发病两周后收获, 收获前测定白粉病发病率、光合指标, 收获时采集不同处理植株相同部位的新鲜叶片, 用于测定生理生化指标。
试验包括轻度、中度和重度3个放牧强度, 各放牧强度包括自然土、灭菌土、无AM真菌土和接种AM真菌土4个土壤处理, 每个处理4个重复。
1.4 收获及指标测定
1.4.1 老芒麦白粉病发病率调查 每盆随机选取20个叶片, 记录发病叶片数。
每盆发病率=发病叶片数/20× 100%。
每个处理4个重复, 各处理下老芒麦白粉病的发病率为4个重复的平均值。
1.4.2 菌根侵染率的测定
染色镜检法测定菌根侵染率[24]:将收获的老芒麦根系冲洗干净后称取0.2 g左右, 置于10% KOH溶液中30 min, 然后于60 ℃水浴中加热1 h, 倾去KOH溶液, 将根系置于1 mol· L-1 HCl溶液中酸化30 s, 倾去HCl; 用蒸馏水冲洗3次后, 用解剖镜检查植物根部皮层中的有色物质是否已除去, 若色素已基本去除, 则用0.05% Trypan Blue染色, 并置于60 ℃水浴保持5~10 min, 冷却后用十字交叉法计算菌根侵染率[25]。
1.4.3 老芒麦总生物量
总生物量:地上、地下部分均先称取鲜重(W1), 然后采用烘干法测定, 先在105 ℃杀青20 min, 然后于80 ℃烘箱烘48 h, 称重(W2)。地上、地下生物量根据测定可溶性糖、AM真菌侵染率等相关指标的鲜重(W3)与干重比, 计算获得。
生物量=
总生物量=地上生物量+地下生物量。
1.4.4 老芒麦磷含量测定
植物地上地下全磷:采用钼锑抗吸光光度法, 建立标准曲线, 用分光光度计(Pectrum, Shanghai 721, 中国), 于700 nm处测定吸光度值[26]。由地上地下全磷含量计算得出总磷含量。
1.4.5 老芒麦叶绿素含量和光合速率
叶绿素含量:采用丙酮浸提法[27]测定, 称取0.1 g新鲜植物叶片, 加丙酮处理获得提取液, 用分光光度计在波长665、645和652 nm下测定吸光度值。
光合速率:早晨09:00-11:00用LI-6400便携式光合测定仪测定植株光合速率。测量时选择生长部位和生长状况一致的叶片进行测定, 每次测定5个重复。
1.4.6 老芒麦防御酶活性
多酚氧化酶(PPO)测定采用分光光度法[27], 于398 nm 波长下测定吸光度。
1.5 数据统计与分析
试验数据录入Microsoft Excel 2007进行整理并计算、作图。用SPSS 19.0统计软件对数据进行显著性检验:分析过程中首先要对数据进行Levenes方差齐性检验, 若方差齐性(P> 0.05), 则可进一步分析。若方差不齐(P< 0.05)则先转换数据, 再进行显著性检验。
2 结果与分析
2.1 老芒麦发病率
所有放牧强度和土壤处理下, 老芒麦均发生了不同程度的白粉病。与轻度放牧相比, 中度和重度放牧白粉病发病率分别降低了19.02%和25.03%(图1)。
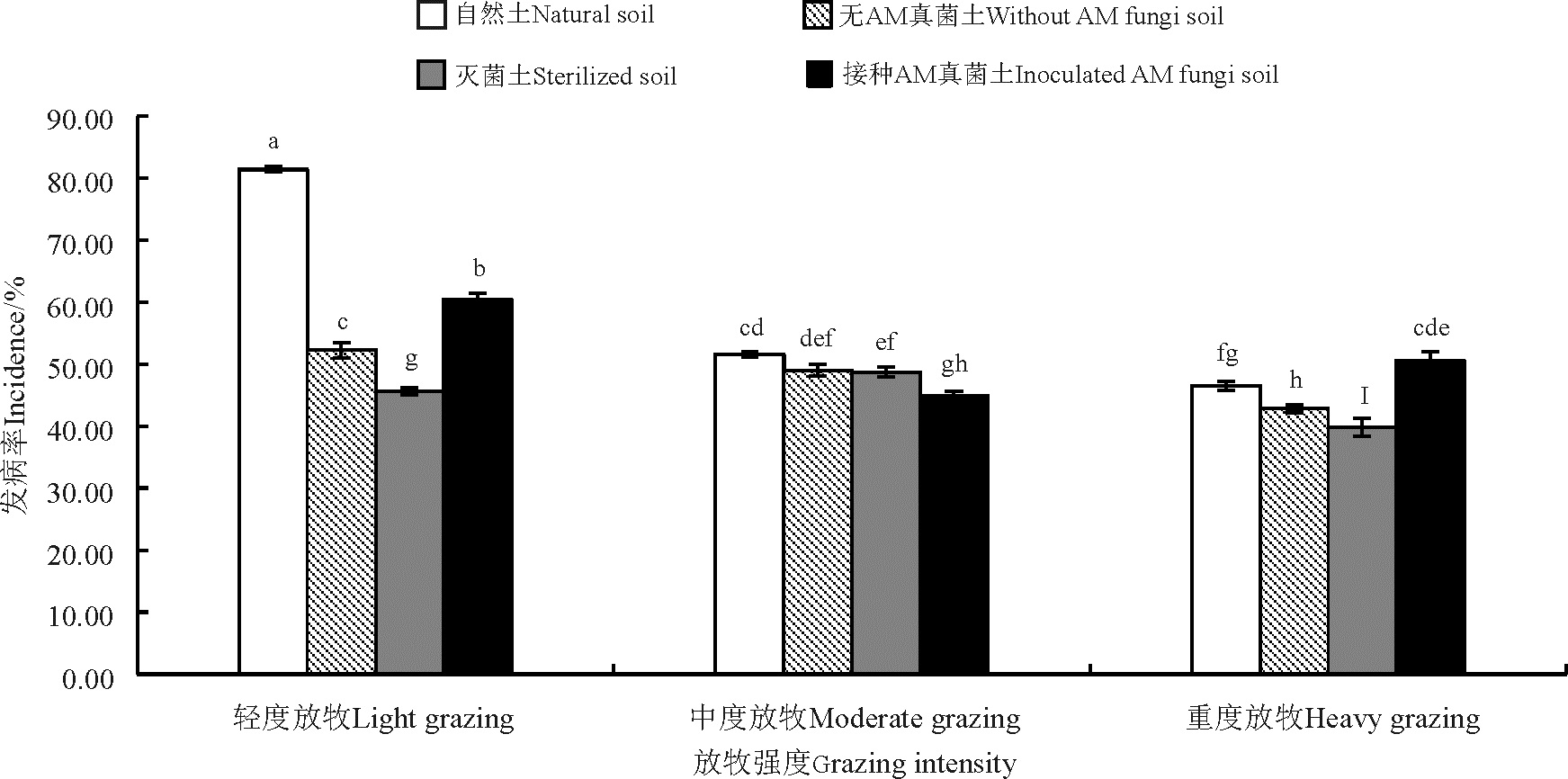 | 图1 不同放牧强度处理下老芒麦白粉病发病率 注:不同小写字母表示不同处理间差异显著(P< 0.05)。下同。Fig. 1 Powdery mildew incidence in Elymus sibiricus under different grazing intensities Note:Different lowercase letters indicate significant difference among different treatments at the 0.05 level. similarly for the following figures. |
各土壤处理, 以自然土处理的老芒麦白粉病发病率最高。与自然土处理相比, 灭菌土、无AM真菌和接种AM真菌处理的老芒麦白粉病发病率分别降低了25.24%、19.75%和13.15%。放牧强度和土壤处理对老芒麦白粉病发病率存在交互效应(P< 0.05), 其中, 以重度放牧的灭菌土处理老芒麦白粉病发病率最低, 为39.84%。
2.2 老芒麦AM真菌侵染率
灭菌土和无AM真菌处理的老芒麦根系未检测到菌根结构。自然土及接种AM真菌的处理老芒麦根系均形成了菌根结构, 且AM真菌侵染率因放牧强度和土壤处理而异(表1)。自然土处理下, 与轻度放牧相比, 中度和重度放牧的老芒麦AM真菌侵染率分别提高了23.93%和39.49%; 接种AM真菌处理下, 与轻度放牧相比, 中度和重度放牧的AM真菌侵染率分别提高了19.57%和48.32%(表1)。
| 表1 不同放牧强度处理下老芒麦AM真菌侵染率 Table 1 AM colonization of Elymus sibiricus under different grazing intensities |
注:同列不同字母表示不同放牧强度间差异显著(P< 0.05)。
Note:Different lowercase letters within the same column indicate significant difference among the grazing treatments at the 0.05 level.
2.3 老芒麦总生物量
放牧可显著影响老芒麦总生物量。与轻度放牧相比, 重度放牧提高了23.46%(图2), 中度放牧下虽有提高, 但差异不显著(P> 0.05)。土壤处理以自然土处理的老芒麦总生物量最低, 显著低于其他处理(P< 0.05)。与自然土处理相比, 无菌土、无AM真菌土和接种AM真菌土处理的老芒麦总生物量分别提高了67.21%、62.89%和56.46%。放牧强度和土壤处理对老芒麦地下生物量不存在交互效应(P> 0.05)。
2.4 老芒麦地上地下总磷含量
放牧、土壤处理对老芒麦地上地下总磷含量的影响与对总生物量的影响较为一致。与轻度放牧相比, 中度和重度放牧分别显著提高了34.02%和33.07%。土壤处理以自然土处理的老芒麦总磷含量最低, 显著低于其他处理(P< 0.05), 与自然土处理相比, 灭菌土、无AM真菌和接种AM真菌处理的老芒麦总磷含量分别提高了186.34%、176.22%和193.09%。不同放牧强度和土壤处理对老芒麦总P含量存在交互效应(P< 0.05)。自然土处理下各放牧强度的老芒麦总磷含量显著低于其它土壤处理, 与自然土处理相比, 灭菌土处理下轻度、中度和重度放牧分别提高了200.85%、285.11%和122.22%; 无AM真菌处理下轻度、中度和重度放牧分别提高了198.1%、285.1%和102.27%; 接种AM真菌处理下轻度、中度和重度放牧分别提高了179.7%、350.42%和111.83%(图3)。
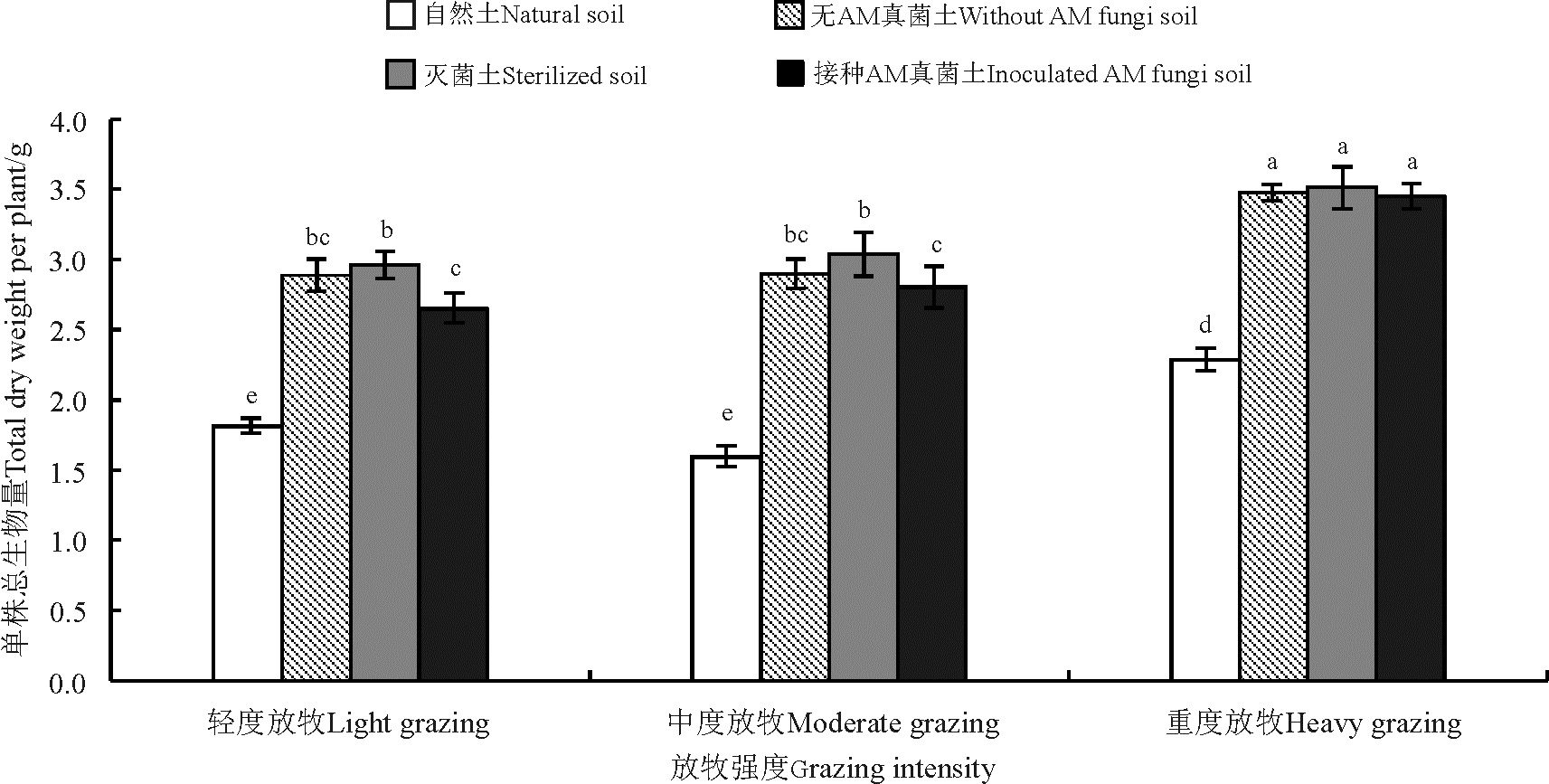 | 图2 不同放牧强度处理下老芒麦总生物量Fig. 2 Total dry weights of Elymus sibiricus under different grazing intensities |
 | 图3 不同放牧强度处理下老芒麦总磷含量Fig. 3 Total P of Elymus sibiricus under different grazing intensities |
2.5 老芒麦叶绿素含量
放牧强度未显著影响老芒麦叶绿素含量(P> 0.05), 与轻度放牧相比, 中度放牧和重度放牧虽提高了老芒麦叶绿素含量, 但差异不显著。土壤处理以自然土处理的老芒麦叶绿素含量最低, 与自然土处理相比, 灭菌土、无AM真菌和接种AM真菌处理分别显著提高老芒麦叶绿素含量40.42%、22.23%和43.28%(P< 0.05)。不同放牧强度和土壤处理对老芒麦叶绿素含量存在交互效应(P< 0.05), 自然土处理下各放牧强度叶绿素含量显著低于其它土壤处理, 其中, 与自然土处理相比, 灭菌土处理下轻度、中度和重度放牧的老芒麦叶绿素含量分别提高了28.29%、40.27%和52.05%; 无AM真菌处理下轻度、中度和重度放牧分别提高了9.39%、27.78%和28.82%; 较接种AM真菌处理下轻度、中度和重度放牧分别提高了65.44%、40.78%和24.78%(图4)。
2.6 光合速率
放牧强度对老芒麦光合速率无显著影响(P> 0.05)。土壤处理以自然土处理老芒麦光合速率最低, 显著低于其他处理(P< 0.05)。与自然土处理相比, 灭菌土、无AM真菌和接种AM真菌处理分别提高了9.85%、69.36%和102.23%。放牧强度和土壤处理对老芒麦光合速率不存在交互效应(P> 0.05)(图5)。
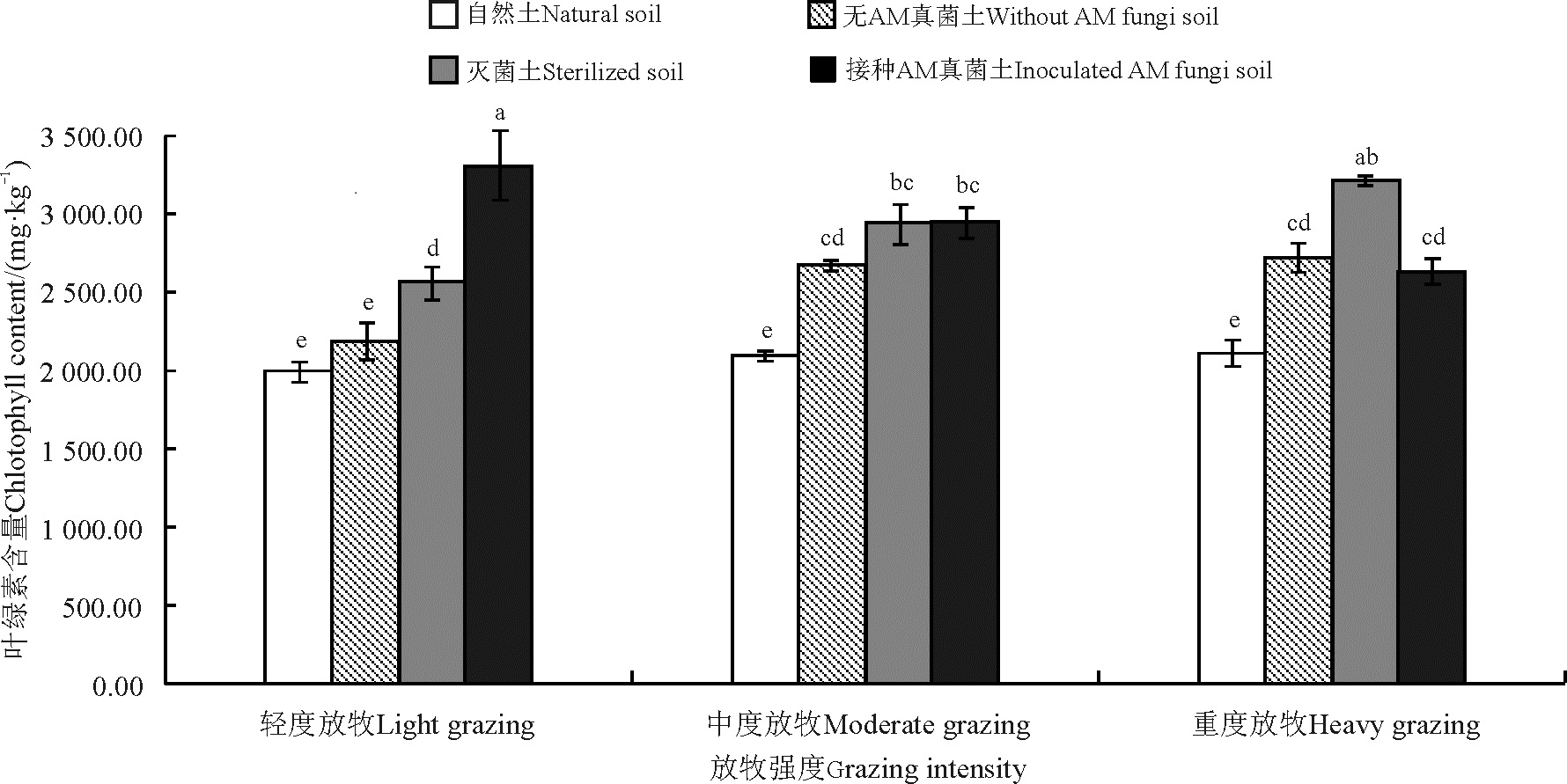 | 图4 不同放牧强度处理下老芒麦叶绿素含量Fig. 4 Chlorophyll contents of Elymus sibiricus under different grazing intensities |
2.7 多酚氧化酶(PPO)活性
放牧强度显著影响老芒麦PPO酶活性(P< 0.05), 与轻度放牧相比, 中度放牧和重度放牧显著降低了42.04%和19.45%。各土壤处理以灭菌土处理下老芒麦PPO酶活性最高, 接种AM真菌处理下老芒麦PPO酶活性最低。与灭菌土处理相比, 自然土、无AM真菌和接种AM真菌处理分别降低了13.0%、12.87%和17.26%。不同放牧强度和不同土壤处理对老芒麦PPO酶活性存在交互效应(P< 0.05), 灭菌土处理下各放牧强度老芒麦PPO酶活性显著高于其它土壤处理。其中, 较自然土处理下轻度、中度和重度放牧分别提高了8.18%、7.47%和22.5%; 较无AM真菌处理分别提高了15.44%、14.33%和8.88%; 较接种AM真菌处理下重度和重度放牧分别提高了35.83%和23.81%(图6)。
 | 图5 不同放牧强度处理下老芒麦光合速率Fig. 5 Photosynthetic rates of Elymus sibiricus under different grazing intensities |
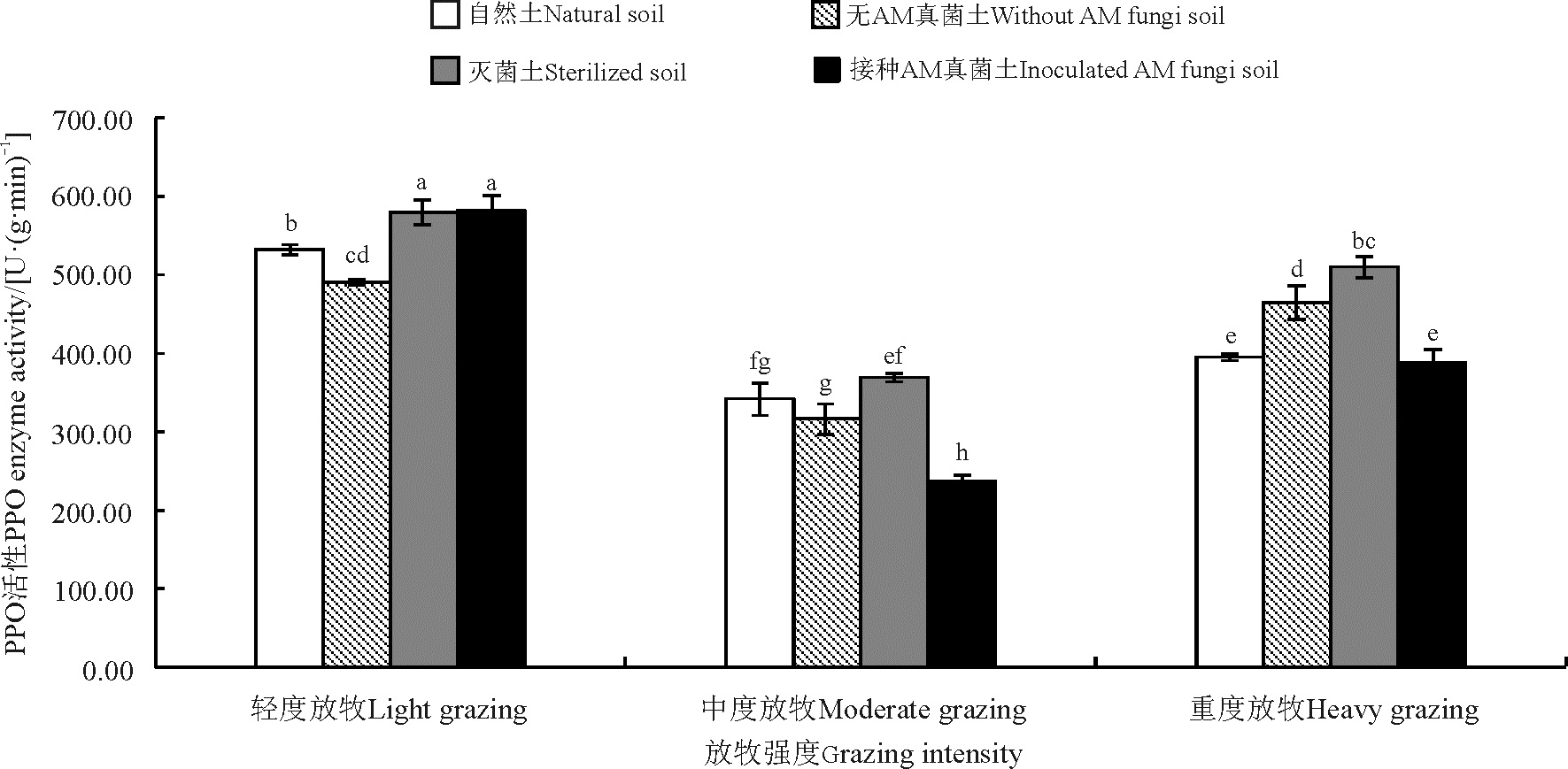 | 图6 不同放牧强度处理下老芒麦PPO酶活性Fig. 6 PPO enzyme activities of Elymus sibiricus under different grazing intensities |
3 讨论与结论
本研究发现, 接种AM真菌与自然土处理下老芒麦根系均形成菌根结构, 接种AM真菌未显著提高AM真菌侵染率, AM真菌侵染率随着放牧强度的增加而增加, 以重度放牧最高, 中度放牧次之, 轻度放牧最低。但通过研究内蒙古草原不同放牧强度下AM真菌的变化, 发现随着放牧强度的增加, AM真菌的侵染率显著降低, 其原因可能为过度放牧时家畜的采食和践踏导致地表植物物种种类减少、盖度下降[16], 从而使植物向地下部输送碳源不能满足大量AM真菌的生长需求[17]。Lugo等[28]报道了放牧对阿根廷天然草地AM真菌侵染率的影响, 结果表明, 放牧未影响AM真菌侵染率。这些结果可能与放牧地土壤养分含量有关。土壤养分含量过高或过低都会降低菌根的侵染率及其在土壤中繁殖的数量, 在一定范围内丛枝菌根真菌的数量随有机质含量的升高而增多, 但当有机质含量超过一定范围, 丛枝菌根真菌的数量就会下降[29]。
不同放牧强度土壤处理下, 老芒麦生物量呈现出重度放牧高于轻度放牧和中度放牧的趋势, 这可能是因为连续放牧的草地, 家畜粪便积累较多, 家畜排泄物提高了放牧地土壤的营养养分含量, 从而促进植物生长, 使植株产量、株高以及生殖枝枝数均提高[30, 31], 从而增加了植物生物量, 增强其抗病性。方爱国和李春杰[32]的研究表明AM真菌可促进野大麦(Hordeum brevisubulatum)养分(P)的吸收, 并显著增加其生物量。本研究表, 老芒麦总生物量灭菌土> 无AM真菌处理> 接种AM真菌处理> 自然土, 老芒麦白粉病发病率自然土> 接种AM真菌处理> 无AM真菌处理> 灭菌土, 说明与不含AM真菌的处理相比, AM真菌的存在反而降低了老芒麦总生物量, 加重了白粉病发病率。这与Duan等[33]的研究结果相似, 可能是因为AM真菌作为共生微生物, 需消耗一定的寄主植物养分, 尤其在植株感病后, 共生微生物与病原菌竞争养分[34], 从而抑制植物生长, 加重病害。
光合作用是衡量植物生长状况的重要指标, 是植物合成有机物质和获得能量的根本所在。而叶绿素是植物进行光合作用不可缺少的, 其含量的多少能影响植物光合作用的强弱。病菌的侵染会破坏叶片绿色组织, 从而使光合作用减弱[35]。本研究发现, 老芒麦在轻度放牧强度下, 接种AM真菌处理的老芒麦叶绿素含量和光合速率均显著高于自然土、灭菌土和无AM真菌处理, 而重度放牧下接种AM真菌处理的老芒麦叶绿素含量低于无AM真菌处理。这可能与放牧过程中家畜散布粪便的养分有关[30, 31]。研究表明, 菌根真菌促进植物生长的效应与菌根侵染改善植物磷营养密切相关, 菌根植物吸收与利用磷的能力显著高于非菌根植物, 尤其在供磷不足土壤环境中, 其作用更加显著[36]。重度放牧条件下, 家畜散布到土壤中的粪便较多, 因而土壤中N、P等养分含量高, 从而抑制了AM真菌的效应, 而轻度放牧条件则相反。
有研究表明, 正常状态下植物体内活性氧代谢处于动态平衡状态的主要原因是有活性氧清除酶系统(SOD、POD、CAT、PPO等)的存在[37], 受到逆境胁迫后, 植物的各个组织结构及其自身的生理生化通常会发生一系列变化, 从而进行主动防卫。寄主的主动防卫常与防御性保护酶活性密切相关。本研究结果表明, 重度放牧提高了老芒麦的POD酶活性。这可能是因为在轻度和中度放牧强度下, 家畜散布的粪便较少, 因而N、P等含量较低, 从而发挥了AM真菌的作用, 使植物产生有毒的次生代谢物来抵抗逆境干扰[38], 而重度放牧条件下则相反。
综上所述, 本研究发现, 放牧和AM真菌均可影响老芒麦生长和白粉病的发生, 与轻度放牧相比, 中度放牧和重度放牧减轻了老芒麦白粉病发病率, 提高了总生物量、磷含量、光合速率; 与自然土相比, 接种AM真菌处理老芒麦白粉病发病率降低了13.15%, 总生物量增加了107.1%。放牧强度和土壤处理对老芒麦叶绿素含量、多酚氧化酶等的影响因放牧、土壤处理组合而异。
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|