


{kind=link}
{kind=link}
{kind=link}
{kind=link}
多枝柽柳幼苗生长及其根系解剖结构对水盐胁迫的响应
[张瑞群1 , 马晓东1, 2  , 吕豪豪
, 吕豪豪1 ]
, 吕豪豪|
|


第一作者:张瑞群(1991-),女,新疆乌鲁木齐人,在读硕士生,主要从事植物生态研究。E-mail:ruiqun1213@126.com
本研究以塔里木河下游本土优势灌木多枝柽柳( Tamarix ramosissima)为研究对象,采用单因素盆栽控制试验,设置不同水分梯度(CK,75% FWC;D1,55% FWC;D2,35% FWC;D3,15% FWC;D4,水淹),不同盐分梯度(CK,基底盐的0.02%;轻盐胁迫S1,基底盐的0.2%;中盐胁迫S2,基底盐的0.5%;中高盐胁迫S3,基底盐的0.8%;高盐胁迫S4,基底盐的1.5%),研究水、盐胁迫对多枝柽柳幼苗生长、根系显微及超微结构的影响。结果表明,1)水分胁迫中,水淹处理60 d后显著增加了幼苗株高、冠幅、生物量的积累( P<0.05),株高相对增长率、冠幅相对增长率分别是CK的1.9和0.9倍。盐胁迫下,随着盐浓度的增高,多枝柽柳幼苗的株高、冠幅相对增长率及生物量减小,S3处理下幼苗生物量显著下降83.42%( P<0.05)。2)盐胁迫下,随着盐浓度的增加根木质部导管孔径降低,S3处理组较对照组显著降低了64.15%( P<0.05),导管密度显著高于CK( P<0.05);水分胁迫下导管孔径和密度呈相反趋势。3)水、盐胁迫下,根细胞超微结构白色体中淀粉粒都有不同程度的肿胀变形,并伴有大量囊泡的产生。盐胁迫下细胞器有解体现象,中高盐胁迫S3下,白色体中淀粉粒膨大,膜破裂解体,细胞核核膜消失,而水分胁迫中未见解体现象,说明盐胁迫对多枝柽柳幼苗的影响大于水分胁迫。
Tamarix ramosissima is a dominate shrub in the lower reaches of the Tarim river. Using a single factor control test with indoor potted plants, five moisture gradients were employed: groups CK, D1, D2, D4, and D5 (flooding), for which soil water contents were 75%, 55%, 35%, 15% field water capacity, and flooding treatment, respectively. For salt stress, the plant were treated with one of five salinity (CK, S1, S2, S3, S4), soil salt contents were 0.02%, 0.2%, 0.5%, 0.8%, 1.5%. We analyzed the effects of water and salt stress on T. ramosissima seedlings by measuring their growth, the roots of microstructure and ultrastructure. The results showed that: 1) under water stress, the flooded D4, promoted the seedling height, crown diameter and biomass accumulation significantly. The relative growth rate of the seedling height and crown width were 1.9 times and 0.9 times of CK, respectively. With the increase of salt concentration, relative growth rates of plant height and crown width increased, and the biomass decreased. In the treatment S3, T. ramosissima seedlings biomass decreased significantly at a rate of 83.42%. 2) Under salt stress, the vessel diameter of the root xylemdecreased, and the diameter of the S3 treatment decreased by 64.5% significantly when compared with that of the control, but vessel densities were higher than that of control group. Opposite trends of the vessel variations were observed under water stress. 3) There were swelling and deformation of starch grains in the root cells under different water and salt stresses, and accompanied by a large number of vesicles secreted. But there was a disintegration phenomenon of organelles under salt stress. Starch grains of the root cell in the leucoplast grew large, the membrane of leucoplast and nucleus disintegrated in the treatment S3. While it was not observed in the disintegration of root organelles under water stress. The results of the experiment proved that salt stress had more influence on T. ramosissima seedlings than water stress.
塔里木河下游地处极端干旱区, 干燥少雨, 蒸发强烈[1], 土壤含盐量较高, 并且由于河水漫溢的影响, 离河道越近, 盐分越高, 植物生长受干旱和盐分胁迫严重[1, 2, 3]。多枝柽柳(Tamarix ramosissima)作为塔里木河下游荒漠河岸林的主要建群种, 具有很强的有性繁殖能力, 种子量丰富, 遇水萌发且萌发率高[4]。但“ 幼苗期” 是多枝柽柳生长发育的“ 脆弱期” , 对土壤条件较敏感。2000年后, 我国实施了塔里木河流域综合治理工程, 生态输水过程中复杂的水盐变化使得河岸边自然和人工漫溢带多枝柽柳幼苗的实时更新受到很大影响, 生态输水对地下水位的抬升发挥了重要作用。输水期间, 下游两岸的河漫滩有利于多枝柽柳种子的萌发, 但大量萌发后的幼苗常会阶段性地处于水淹状态。输水期过后, 上游河水携带的盐分聚集在土壤表层, 干旱区强烈蒸发的作用使盐分不断浓缩, 致使塔里木河下游表层土壤盐分极高[2, 3]。多枝柽柳幼苗的根系分布在浅表层, 深受土壤干旱及盐胁迫的影响, 不利于幼苗后期生长发育。因此, 水分和盐胁迫成为影响多枝柽柳幼苗成活和后期生长繁殖的关键因子[5]。
目前, 对柽柳属植物的研究主要集中在生长发育、生理生化调控、形态学的进化适应等方面[6, 7, 8], 形态解剖特别是盐、干旱胁迫解剖结构特征的响应研究较少。多枝柽柳的根系生长具有很高的表型可塑性[9], 尤其是多枝柽柳根系微观结构变化程度与抗逆性强弱之间的相关性尚不清楚[9, 10], 本研究通过对水分和盐分胁迫后多枝柽柳幼苗根系显微解剖结构及超微结构的变化进行分析, 进一步了解水、盐胁迫对柽柳幼苗根系显微结构的胁迫效应, 揭示水、盐胁迫下多枝柽柳幼苗结构解剖学的抗逆性特征, 为其对生境胁迫的响应提供细胞学证据, 进一步完善对该物种抗逆机理的认识。
2014年4月于塔里木河下游荒漠河岸林采集多枝柽柳种子, 挑选成熟饱满的种子, 风干后将种子放在布袋中, 4 ℃低温储藏, 防止种子失活[11]。同年5月, 将种子播种于25 cm(口径)× 30 cm(高)的花盆中培育。栽培基质为塔里木河下游的河岸沙壤土, 使用前用蒸馏水淋洗除盐, 淋洗后土壤含盐量低于0.02%, 沙壤土最大田间持水量为24.11%, 晒干备用。花盆被放置于玻璃温室受自然光照, 温室内试验期间昼夜平均温度20.9~28.6 ℃、平均湿度24.9%~30.7%。待播种3个月后, 选取长势一致的幼苗进行水、盐胁迫, 每盆定苗10棵, 每个处理6个重复。
水分胁迫设置对照(CK, 土壤含水量为田间最大持水量的75%)、轻度胁迫(D1, 土壤含水量为田间最大持水量的55%)、中度胁迫(D2, 土壤含水量为田间最大持水量的35%)、重度胁迫(D3, 土壤含水量为田间最大持水量的15%)和水淹处理(D4, 仅地下根系部分完全淹没, 茎叶保持干燥)5个水分梯度, 采用称重法进行控水处理, 每天称重保持土壤水分, 共培养60 d。
盐胁迫按含盐量(土壤盐分/土壤干重× 100%)的计算方式配制不同梯度的NaCl溶液来控制。设对照组(CK, 不加盐, 土壤含盐量为基底盐的0.02%)、轻盐胁迫(S1, 土壤含盐量为基底盐的0.2%)、中盐胁迫(S2, 为基底盐的0.5%)、中高盐胁迫(S3, 为基底盐的0.8%)和高盐胁迫(S4, 为基底盐的1.5%)5个处理水平。为避免盐激, 采用少量多次的浇灌方式来控制, 每天递增0.2%的盐分, 直至预定浓度。之后每天称重浇水, 浇水量达到最大持水量的75%, 达到预定浓度后, 培养60 d, 为防止盐分流失, 每个花盆下放一托盘, 当有水渗出花盆时, 将渗漏水倒回花盆, 以保持土壤含盐量不变。
1.2.1 幼苗的生长指标 室内盐分和水分胁迫控制试验开始前按处理水平分别测量幼苗的株高和冠幅, 在试验结束后立即测量不同处理下幼苗的株高, 冠幅和地上、地下生物量, 每个处理6个重复。相关指标按以下公式计算:
相对株高增长率=(最终植物高度-最初植物高度)/最初植物高度× 100%;
相对冠幅增长率=(最终植物冠幅-最初植物冠幅)/最初植物冠幅× 100%;
根冠比=地下部分生物量/地上部分生物量。
1.2.2 幼苗根解剖结构的观测 在试验结束时进行毁灭性取样, 选取形态学下端幼苗的主根中段2/3的部分, 截成1 cm的小段, 取3个重复放入70%的FAA 固定液中固定, 采用常规石蜡切片法制片[12]。待装片风干后, 用 MOTIC数据显微图像系统软件测量表皮厚度、皮层厚度、导管孔径、导管密度等指标, 每个装片记录15个视野, 计算平均值。除导管密度外, 以占径比来表示不同组织厚度在整个器官中所占的比例[13]。组织占径比=组织厚度/根直径厚度。
超微结构的观察, 将主根形态学下端2/3部分的根切割成1 mm的小段, 浸入35%的戊二醛固定液中放置24 h, 同时抽气直到切块下沉为止。固定后经过常规透射电镜处理方法处理后[14], 在日立 H-600型透射电镜下观察, 照相。
所得生长指标及根解剖结构指标的数据均采用SPSS 19.0进行单因素方差分析, 并用Duncan法对数据进行多重比较; 并采用Excel 2007制图。
水分胁迫1周后, 重度胁迫D3处理下的幼苗全部死亡(图1)。水分胁迫处理60 d结束后, 幼苗株高相对增长率呈CK> D1> D2的趋势(P> 0.05), 水淹处理D4的幼苗株高相对增长率显著增高, 为CK的1.9倍(P< 0.05)。幼苗冠幅相对增长率依次为D1> D2> CK(P> 0.05), 水淹处理D4的冠幅相对增长率比CK高90.28%(P< 0.05)。幼苗单株生物量在中度胁迫D2下最小, 仅为0.08 g; 水淹处理D4的单株生物量最大, 为0.33 g, 显著高于CK(P< 0.05), 是CK的165%。幼苗根冠比在轻度胁迫D1下最大, 显著高于CK(P< 0.05), 中度胁迫和水淹处理的根冠比高于CK, 但差异不显著(P> 0.05)。水淹处理下, 株高和冠幅的显著增大, 可以提高同化枝及叶的光合作用, 有助于提供充足的O2和C。
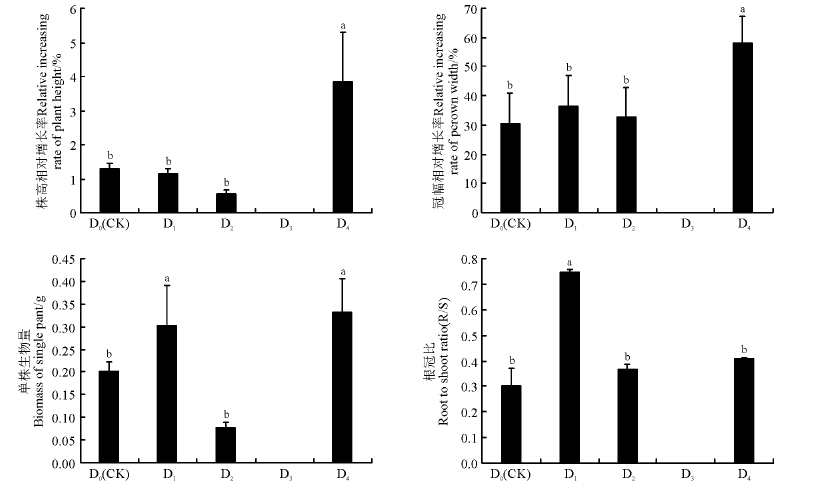 | 图1 水分胁迫对多枝柽柳幼苗生长的影响Fig.1 Effects of water stress on growth of the Tamarix ramosissima 注:D0(CK)、D1、D2、D3和D4分别指土壤水分含量为田间最大持水量(FWC)的75%、55%、35%、15%和水淹处理。表1、图3同。 Note: D0(CK), D1, D2, D3, and D4 means soil moisture are 75%, 55%, 35%, 15% of field water content (FWC) and flooding treatment, respectively. The same in Table 1 and Fig.3 |
随盐浓度的升高, 多枝柽柳幼苗的生长状况较对照均变差。高盐胁迫S4处理下, 胁迫3周内幼苗连续干枯死亡, 至第5周全部死亡。S2和S3中盐胁迫1周后, 幼苗出现叶片变黄现象, 胁迫后期变黄情况加重, 并出现死苗状况。盐胁迫条件下, 幼苗的株高相对增长率、冠幅相对增长率以及单株生物量均显著低于CK(P< 0.05)(图2)。相较于CK, 中高盐胁迫S3下多枝柽柳幼苗单株生物量显著下降(P< 0.05), 下降幅度达70%。根冠比随盐浓度的增加呈先升高后降低的趋势, 中盐胁迫S2下根冠比达到最大, 显著高于CK(P< 0.05)。
 | 图2 盐胁迫对多枝柽柳幼苗生长的影响Fig.2 Effects of salt stress on growth of the Tamarix ramosissima 注:S0(CK)、S1、S2、S3和S4分别指土壤含盐量为基底盐的0.02%、0.2%、0.5%、0.8%和1.5%。表2、图4同。 Note: S0(CK), S1, S2, S3 and S4 means soil salt contents were 0.02%, 0.2%, 0.5%, 0.8%, and 1.5% of soil base salt content, respectively. The same in Table 2 and Fig.4. |
水分胁迫对多枝柽柳幼苗根解剖结构的影响显著(表1)。随着水分胁迫加剧, 给水量减少, 幼苗根部表皮厚度占径比逐渐增大, 均显著高于对照(P< 0.05); 幼苗根皮层厚度占径比也增大, 且中度干旱胁迫D2与CK相比显著增加了12.96个百分点, 而根维管柱占径比显著下降了14.30个百分点。水淹胁迫D4下幼苗根部维管柱直径降低了50.71%, 与对照相比差异显著。D2和D4处理组幼苗根木质部导管孔径占径比与CK相比显著增大, 在中度胁迫D2下增幅为56.32%。D1和D2处理组及水淹处理组D4幼苗根部导管密度均呈降低趋势, 重度胁迫D3在试验结束时已全部死亡。
| 表1 水分胁迫对多枝柽柳幼苗根系解剖结构的影响(平均值± 标准误差) Table 1 Effect of drought stress on shoot anatomical characteristics of the Tamarix ramosissima(mean± SE) |
随着盐浓度的增加, 多枝柽柳幼苗根表皮厚度占径比增大且皮层薄壁细胞排列更紧密, 中高盐胁迫S3与CK相比, 增加了52.85%(P> 0.05)(表2)。根皮层厚度占径比随盐胁迫加剧而降低, S2处理组显著低于CK(P< 0.05); 根维管柱厚度占径比则逐渐增加, 且在中高盐胁迫S3下维管柱占径比与CK相比显著增加5.07个百分点(P< 0.05)。但随着盐浓度增加, 根维管柱木质部中导管孔径占径比显著减小, S3与对照相比显著降低64.16%。与导管孔径变化趋势相反, 导管密度随盐胁迫加剧而显著增大。高盐胁迫S4处理下幼苗在试验结束时已全部死亡。
| 表2 盐胁迫对多枝柽柳幼苗根系解剖结构的影响(平均值± 标准误差) Table 2 Effect of salt stress on shoot anatomical characteristics of the Tamarix ramosissima(mean± SE) |
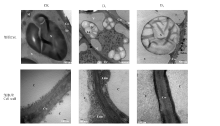
对照组多枝柽柳幼苗的根细胞中细胞膜结构清晰, 白色体结构完整(图3)。在中度干旱胁迫D2中, 根细胞白色体及淀粉粒数量增多, 淀粉粒膨大占据整个白色体, 细胞质膜内陷形成大小不一的囊泡; 细胞核膜结构完整, 结构清晰, 但出现核固缩现象。干旱胁迫下根细胞壁均有变薄现象。水淹处理下, 白色体膨大解体, 细胞质膜活跃, 内陷成大小不一的囊泡, 有些内陷的囊泡中含有高电子致密物质; 水淹处理下根细胞细胞壁与对照相比增厚。
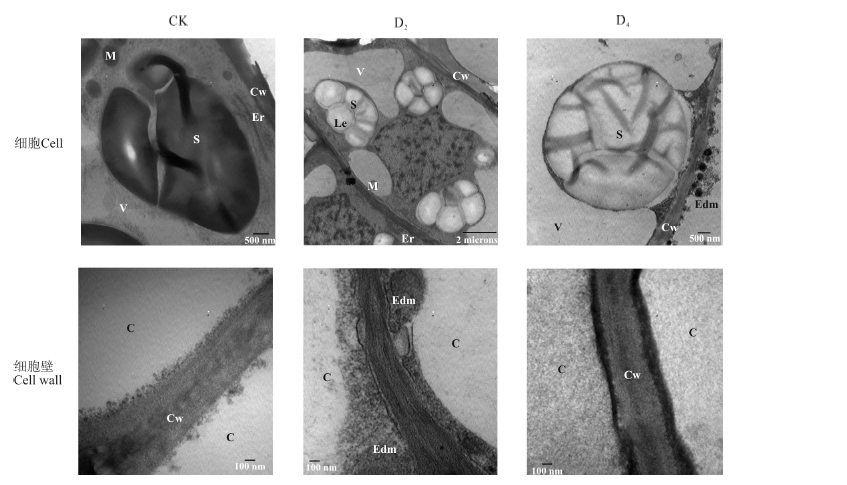 | 图3 水分胁迫对多枝柽柳根细胞超微结构的影响Fig.3 Effect of water stress on the root ultrastructure of Tamarix ramosissima 注:C, 细胞; Cw, 细胞壁; M, 线粒体; N, 细胞核; S, 淀粉粒; V, 液泡; Le, 白色体; Lem, 白色体被膜; Edm, 电子致密物质。图4同。 Note: C, cell; Cw, cell wall; M, mitochondria; N, cell nucleus; S, starch grains; V, vacuole; Le, leucoplast; Lem, leucoplast envelop; Edm, electron-dense material. The same in Fig.4. |
随着盐浓度的增大, 幼苗根细胞变化明显。轻度盐胁迫S1下即引起幼苗根系细胞中白色体肿胀变形, 并含有较多大的淀粉粒(图4)。随着胁迫程度的增加, 中盐胁迫S2下细胞核结构完整, 但核膜模糊甚至消失; 中高盐胁迫S3下, 淀粉粒膨大, 使得白色体膜结构模糊破损, 最终解体。随着盐胁迫程度的增加细胞质膜内陷为许多囊泡, 含有高电子致密物质, 有的囊泡延伸至其它细胞器, 如白色体。盐胁迫处理组的细胞壁厚度高于CK, 且高于干旱胁迫处理组。
 | 图4 盐胁迫对多枝柽柳根细胞超微结构的影响Fig.4 Effect of salt stress on the root ultrastructure of Tamarix ramosissima |
根系直接接触土壤, 是植物吸收水分和养分的主要器官, 所以土壤逆境对植物的危害最主要的部位就是根系[16]。研究发现, 水、盐胁迫处理试验中, CK的多枝柽柳幼苗长势最好, 水分胁迫中的干旱胁迫和盐胁迫都不利于其生长。水、盐胁迫下, 根系首先感应并发出信号, 同时根系形态结构、生理化学、生物量等也发生相应变化, 进而抑制植物生长, 甚至导致植物死亡[17]。研究表明, 在逆境下, 植物能够感应外界胁迫, 在长期进化的过程中形成了一系列抵御不利环境的机制, 如植物体外部形态和细胞形状结构的改变, 生理生化代谢途径的适应等[16]。根系感知与响应胁迫的能力在很大程度上依赖于其对不断变化的土壤环境的适应[18]。本研究中, 水淹处理模拟了塔里木河下游短暂的河水漫溢过程, 水淹使幼苗地上部分的相对生长率、生物量显著增加, 但根冠比并没有与之同步增大。因此, 本研究认为, 在随后而来的干旱期, 水淹后的幼苗的根系生长状况对它后期生存不利。
本研究中, 水分胁迫下多枝柽柳幼苗根表皮和皮层厚度占径比增加, 中度干旱胁迫D2处理下皮层厚度占径比与CK相比显著增加了12.96个百分点, 这有利于根系储存水分。随着多枝柽柳幼苗的生长, 处理组导管孔径占径比显著高于CK, 中度干旱胁迫D2与CK相比显著增大了56.32%, 导管孔径增加提高了木质部导管单位面积的输水量, 提高多枝柽柳幼苗的抗旱性[18, 19]。水分胁迫时, 达乌里胡枝子(Lespedeza davurica)导管比正常供水发达, 其导管直径变大对水分的输导更有利, 增强了抵抗水分的能力[20]。水淹处理下的表皮、皮层占径比显著高于对照组, 增幅分别为57.38%、33.86%, 同时表皮和皮层栓质化明显。这与杨朝东等[21]的研究结果类似, 他们认为栓质化的表皮皮层形成了质外体屏障, 阻挡了水和离子的自由进出, 起到保护植物生理功能的作用。本研究发现, 水淹处理下多枝柽柳根组织中薄壁细胞碎裂生成通气组织, 以供给植物氧气。通气组织是根部有氧呼吸便利的通道, 可以保证植物受到较少伤害[22]。随着塔里木河下游生态输水的结束, 河岸漫溢带的水淹地带随着该地区强大的蒸发量, 又会对幼苗产生干旱胁迫。而此时幼苗根薄壁细胞已经不完整, 碎裂生成通气组织, 维管柱占径比降低, 面对干旱的逆境, 我们推测解剖结构的这种变化不利于幼苗在塔里木河下游输水结束后干旱时期的生存。
盐胁迫下, 植物主要通过稀盐、泌盐、拒盐3种方式来减少盐对机体的损害, 例如, 液泡区域化、盐腺或盐囊泡的泌盐、根细胞膜对盐离子的选择透过性、凯氏带阻挡等方式[23, 24]。盐胁迫导致土壤水势降低, 植物受到水分胁迫及盐害双重胁迫。多枝柽柳通过根系拒盐和位于同化枝及叶表面的盐腺泌盐, 以减轻盐胁迫的危害[24]。柽柳通过根系拒盐可以将90%到达根表面的Na+阻挡在根外, 将进入植物地上部约50%的Na+分泌掉, 说明根系拒盐是柽柳最主要的抗盐机制[25]。本研究中, 盐胁迫下, 根组织表皮细胞厚度增大, 占径比增幅为52.85%, 有效阻挡了盐离子的进入。导管孔径占径比与水分胁迫相比变化趋势相反, 幼苗皮层厚度占径比下降幅度达19.81%, 这有助于缩短根部水分吸收距离, 维管柱占径比增加, 提高疏导水分的能力。有学者指出, 盐生植物根组织发达的表皮和外皮层能够大大抑制有害离子进入根细胞, 能够有效缓解盐害[10]。还有研究表明, 盐胁迫下植物根系内皮层细胞的栓质化能够降低Na+向维管柱转移[26]。本研究中, 皮层厚度占径比呈降低趋势, 但通过观察解剖结构发现也有栓质化现象。玉米(Zea mays)通过根结构中维管柱的加厚、木质化程度加深, 增加导管数量, 以提高根的运输能力, 适应低K+胁迫[27]。这与本研究结果类似, 本研究中木质部中导管孔径占径比显著低于CK, 降幅达64.15%; 木质部导管密度却高于CK, 这可能是多枝柽柳幼苗通过调节导管来调节控制盐分进入体内的一种方式。
根细胞的超微结构在逆境下发生了变化, 盐胁迫下, 高粱(Sorghum vulgare)根的表皮和皮层细胞中液泡数量均显著增加[28]。本研究中, 盐胁迫下根细胞壁厚度高于CK和水分胁迫处理组。Cd2+胁迫下, 玉米根尖细胞超微结构膜系统破坏明显, 细胞膜、核膜、线粒体、高尔基体等膜结构模糊不清, 最终细胞器消失, 细胞解体[29]。当离子聚集到一定程度, 植物细胞本身的盐胁迫耐受力不足时, 细胞及细胞膜系统遭到破坏, 细胞膜内陷成大量囊泡, 这些囊泡中的高电子致密物质含有NaCl, 以减少细胞质中游离的盐离子, 减少对细胞的损害[30]。白色体与叶绿体一样, 都是从前质体发育而来, 但是对白色体的研究较少, 对叶绿体的研究表明, 盐胁迫下的植物叶绿体内淀粉粒数目显著增多, 体积增大[31]。本研究与其类似, 水、盐胁迫下白色体中的淀粉粒数目增多, 体积膨大, 以维持多枝柽柳幼苗在逆境下的正常生长, 既缓解能量的短缺, 保证细胞正常的生命活动, 又提高了细胞的渗透压, 有利于水分的吸收与保持[25]。水、盐胁迫下, 细胞核均出现核膜模糊, 核固缩的现象。作为细胞系统的调控中心, 对细胞核的破坏意味着细胞生命活动的紊乱, 最终导致细胞死亡[32]。可见, 逆境对多枝柽柳幼苗的胁迫重要机制是对细胞超微结构产生不可逆的损伤, 破坏了细胞正常的生命活动。
在逆境条件下, 植物体内的水分和盐分变化直接影响到根系的细胞结构, 并严重影响到植物对水分和营养物质的吸收, 最终表现在植物的表型上。细胞数目的增多及个体的长大才能促进植物的生长[28], 轻度水分胁迫下多枝柽柳幼苗的生长并未受到影响, 而中度水分胁迫及盐胁迫下幼苗的生长明显受到抑制, 主要是由于根细胞中细胞核受损, 破坏了细胞的正常生命活动, 导致植物细胞分裂出现障碍。
综上所述, 水、盐胁迫对多枝柽柳幼苗的生长均有一定影响。幼苗通过调整生长、生物量投入、形态解剖结构的变化来适应水、盐胁迫, 并对水分胁迫有较高的耐受性。多枝柽柳幼苗可通过不断调节根系表皮、皮层厚度以及导管孔径、密度等来适应土壤水分和盐分的变化。水、盐胁迫下多枝柽柳幼苗根系细胞都伴随大量囊泡的产生, 中、重度盐胁迫下细胞核核膜消失, 与水分胁迫相比, 盐胁迫是决定多枝柽柳幼苗生长更为重要的生态因子。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|