


{kind=link}
{kind=link}
{kind=link}
15个白三叶品种的ISSR和ITS遗传多样性分析
[李金博1, 2, 3 , 高丽华2 , 周美亮2 , 李诗刚3 , 彭昭良3 , 吴燕民2  ]
]
]
|
|


第一作者:李金博(1985-),男,河南宜阳人,在读博士生,研究方向为草业科学。 E-mail:jinbo406@126.com
利用ISSR标记和ITS序列分析在我国大面积推广应用的15个白三叶( Trifolium repens)品种的遗传多样性与地理来源关系,试图为我国白三叶新品种的选育提供遗传理论支持。结果表明,15个白三叶品种明显分为两大类,第Ⅰ类有10个白三叶品种,主要来源于丹麦、澳大利亚、新西兰和荷兰;第Ⅱ类有5个白三叶品种,主要来源于美国、中国和阿根廷。白三叶品种聚类具有一定的地域性分布规律,ISSR和ITS均能有效地对白三叶品种进行遗传多样性分析,且ISSR可以更加细致地对其进行分类。
The relationship of genetic diversity with geographical origin between 15 white clover cultivars on the popularization and application of the large area in China, which provides genetic theory support the white clover breeding. The results showed that 15 white clover cultivars were divided into two categories, the class Ⅰ has 10 cultivars of white clover, mainly comes from Denmark, Australia, New Zealand and the Netherlands; There are five white clover cultivars in the class Ⅱ, mainly comes from USA, China and Argentina. White clover cultivars clustering has certain regional distribution, the genetic diversity of white clover cultivars could be studied by using ISSR and ITS analyzing , and ISSR can be more detailed on the classification.
白三叶(Trifolium repens)是豆科(Leguminosae)三叶草属多年生草本植物, 目前在温带、亚热带地区广为种植[1, 2]。白三叶具有茎叶丰富、草质细软、营养价值高、适口性好和产量高的特点, 是优良的豆科牧草, 覆盖的白三叶降解也可以提高果园土壤肥力[3]。同时, 白三叶作为园林地被植物具有观赏价值高、再生能力强、栽植成本低等特点, 在园林绿化上应用越来越多。白三叶在我国大体分布于22° -51° N, 80° -130° E地区, 主要为四川、云南、贵州等省, 在其它省(区)也有分布[4, 5]。白三叶是异花授粉、自交不亲和植物, 自然选择及人工育种得到的品种均具有很强的基因杂合性[6], 而白三叶的遗传多样性和多倍性也影响了白三叶的遗传研究及新品种的培育[2]。
ISSR(inter-simple sequence repeat)分子标记是由Zietkiewicz等提出的一种新型分子标记, 是在SSR(simple sequence repeat)标记基础上发展起来的一种通过半随机性引物进行PCR扩增的技术, 可用于检测重复序列之间区域的DNA序列差异[7], 已广泛应用于多种农作物、果树以及部分草资源的遗传多样性分析、品种鉴定等研究中[8]。利用ISSR分子标记可以对白三叶进行鉴定, 明确其遗传差异和亲缘关系[9]。内转录间隔区(internal transcribed spacer, ITS)位于18S和26S rRNA基因之间, 被5.8S rRNA基因分为两段(ITS1和ITS2)。ITS序列变异较快, 可以提供丰富的变异位点和信息位点, 是许多植物类群系统分类与进化分析的重要分子标记[10]。
近年来, 国内外学者通过形态学分类、同工酶生化标记、DNA分子标记等对白三叶遗传多样性研究已经有一定报道。如, 利用形态学标记和聚类分析方法探究白三叶的起源关系[11]; 利用过氧化物酶与酯酶同工酶检测勒代诺白三叶和其它白三叶品种的同源性[12]; 利用RAPD分子标记分析近缘杂交的白三叶的遗传多样性[13]; 利用AFLP对52个野生及栽培种白三叶种质材料进行聚类分析[14]。通过形态学指标和聚类分析13份新疆野生白三叶的遗传多样性进行分析[15]。国内遗传多样性研究大多采用形态学指标或者同工酶法等, 研究对象也往往是收集的多个野生材料, 而目前生产上常用的白三叶品种相关研究相对较少, 急需科研工作者对其进行研究, 为我国白三叶的生产应用及培育提供理论和实践指导。鉴于此, 本研究利用ISSR和ITS分子标记相互验证, 对目前在我国大面积推广应用的白三叶品种进行遗传多样性分析, 以期阐明白三叶遗传多样性与地理来源之间的关系, 同时为我国白三叶新品种选育提供遗传理论支持。
本研究共选用常用白三叶品种15个(表1), 均于2012年从种子公司购买, 全部种植在北京市顺义区中国农科院试验基地。试验地地处40° 13' N, 116° 38' E, 海拔35 m, 年均降水量约625 mm。土壤为砂壤土, 有机质含量为1.35%, 全氮含量为0.9%, 速效磷含量26.16 mg· kg-1, 速效钾含量42.35 mg· kg-1, 属微碱性土壤, pH为8.3。播种3个月后对植株幼嫩叶片进行取样。
| 表1 供试白三叶品种名称及来源 Table 1 Names and origins of tested white clovers |
DNA提取利用全式金试剂盒(Transgen, China), 每个材料取15株幼嫩叶片混合取样。PCR所需引物均由北京奥科鼎盛生物科技有限公司(AuGCT. Biotech Co., Ltd.)合成。
根据哥伦比亚大学公布的ISSR序列合成100条ISSR引物。ISSR体系为25 μ L, 其中DNA(40 ng· μ L-1)1 μ L , 引物(5 mmol· L-1)1 μ L, 10× buffer 2.5 μ L, MgCl2(25 mmol· L-1)2 μ L, Taq DNA polymerase (1 U)1 μ L, dNTP(10 mmol· L-1)2 μ L和无核酸酶水15.5 μ L。PCR程序为 94 ℃ 5 min; 94 ℃变性30 s, 退火温度 55~59 ℃, 1 min, 72 ℃延伸90 s, 35个循环; 72 ℃延伸5 min。扩增后产物保存在4 ℃冰箱。
电泳检测:1.5%的琼脂糖凝胶, 指示剂为北京原平皓公司生产的gelsafe(10 000× )。采用北京六一厂DYY-7C型电泳仪和DYC-34A型电泳槽在90 V的电压下进行1 h电泳, 电泳液为TAE buffer, 100 bp的DNA Marker。利用伯乐公司凝胶成像系统进行紫外观测, 记录多态性条带。
利用通用引物T4(GGAAGTAAAAGTCGTAACAAGG)和T5(TCCTCCGCTTATTGATATGC)进行扩增。PCR反应体系为25 μ L, DNA(40 ng· μ L-1)1 μ L, 上下游引物各(5 mmol· L-1)1 μ L, 10× buffer 2.5 μ L, MgCl2(25 mmol· L-1)2 μ L, Taq DNA polymerase (1 U)1 μ L, dNTP (10 mmol· L-1) 2 μ L和无核酸酶水14.5 μ L。PCR反应程序:94 ℃预变性2 min; 94 ℃变性30 s, 52 ℃退火30 s, 72 ℃延伸90 s, 共35个循环; 最后72 ℃延伸10 min, 产物于4 ℃保存。PCR产物检测:PCR产物5 μ L用gelsafe染色, 125 V的电压下进行25 min电泳, 经1%的琼脂糖凝胶电泳检测, 用凝胶成像仪照相。
测序:PCR产物经纯化试剂盒(全式金试剂盒, Transgen, China)纯化后送由中国农业科学院重大工程楼进行正反向测序。
ISSR分析根据条带的有无分别标记为1或0。材料间遗传相似系数(GS)根据公式GS=2Nij(Ni+Nj)计算, 其中:Nij为材料Ni和Nj共有的扩增片段数目, Ni为材料i中出现的扩增片段数目, Nj为材料j中出现的扩增片段数目。将GS值按不加权成对群算术平均法(UPGMA)进行遗传相似性聚类。统计分析在NTSYS 2.1软件下进行。
将所有测得的序列登录到Genbank, 获取序列登录号。ITS分析用DNAStar分析各序列同源性, 用CLustalX 2.0完成DNA序列的完全对比, 保存为.aln格式, 然后运用系统发育软件MEGA 4.1软件转换成.meg格式, 系统树以苜蓿(Medicago sativa)(AF053142.1)为外群采用邻接距离矩阵法(neighbor-joining)构建, 应用自展法(bootstrap)进行可信度检测, 重复1 000次。
利用两个白三叶品种海法和胡以阿对100条ISSR引物进行筛选, 从中筛选出条带清晰、重复性高、表达稳定的5条ISSR引物, 即810、816、822、826和836(表2)。
利用所选的5条引物对15个白三叶品种进行PCR正式扩增, 共得到DNA条带41条, 扩增条带变幅为7~10条, 平均每个引物可以扩增出8.2条(表2)。在41条带中, 多态性条带为38条, 扩增多态性条带变幅为6~9条, 平均占比为92.68%, 平均每个引物可以扩增出多态性条带7.6条。这说明ISSR标记能够揭示出白三叶品种之间较高的多态性。
| 表2 ISSR引物序列和特异性条带数目 Table 2 List of ISSR primers and polymorphic bands |
基于遗传相似系数, 对白三叶进行UPGMA聚类分析, 将15个白三叶品种聚为两大类(图1)。第Ⅰ 类中, 丹麦的百事和米尔卡亲缘关系较近, 且这两个品种均为中叶型白三叶; 与小叶型的瑞文德和大叶型的克朗德共同构成了第1亚类; 澳大利亚的荷兰白和考拉亲缘关系较近, 新西兰的胡以阿和铺地关系较近, 以上4个与澳大利亚的海发聚在一起为第2亚类, 其中除铺地为小叶型外, 其它3个品种都为中叶型; 而来自荷兰的雷托单独为第3亚类。说明这些白三叶品种的亲缘远近主要还是与地理区域及引种历史相关。
 | 图1 基于ISSR的15个白三叶品种聚类结果Fig.1 Cluster analysis of 15 white clover cultivars by ISSR |
来自美国的瑞加与来自中国的鄂牧一号聚在一起后, 再与来自美国的拉迪诺、路易斯安那, 以及来自阿根廷的碧盛聚在一起形成第Ⅱ 大类。而鄂牧一号是以瑞加为原始材料选育而来, 聚类分析也证明这两个品种亲缘关系较近。美国与阿根廷均处于美洲大陆, 亲缘关系相对较近。
基于遗传相似系数, 对15个白三叶品种进行主成分分析, 位置相靠近者遗传距离较近(图2)。结果表明, 主成分分析结果和遗传聚类结果类似, 主成分分析结果更直观表明了15个白三叶品种之间的遗传关系。
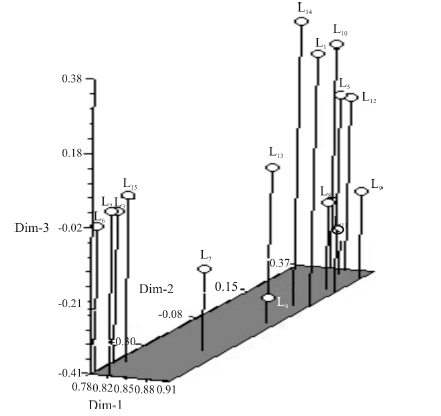 | 图2 基于ISSR的15个白三叶品种主成分分析Fig.2 The principal-coordinate analysis of 15 white clover cultivars by ISSR |
测序结果得到ITS序列扩增长度从671~747 bp, 通过Genbank在线提交所获得的序列, 待审核完成后获得了NCBI中唯一的序列号, L1-L13登陆号为(KR914569-KR914581), L14-L15登陆号为(KR914586-KR914587)。
利用邻接法(NJ)构建系统进化发育树(图3), 可以将15个白三叶品种和苜蓿明显分为三大类。第Ⅰ 类有10个白三叶品种, 包含有来自丹麦的克朗德、米尔卡、百事和瑞文德, 同时也包含了澳大利亚的荷兰白、海法和考拉, 以及新西兰的胡以阿和铺地, 荷兰的雷托, 其中荷兰白和考拉亲缘关系较近; 第Ⅱ 类有5个白三叶品种, 包含有美国的拉迪诺、路易斯安那和瑞加, 同时也包含了中国的鄂牧一号和阿根廷的碧盛。苜蓿单独聚为第Ⅲ 类, 说明本研究结果可靠。ITS聚类分析结果与ISSR聚类分析结果类似。
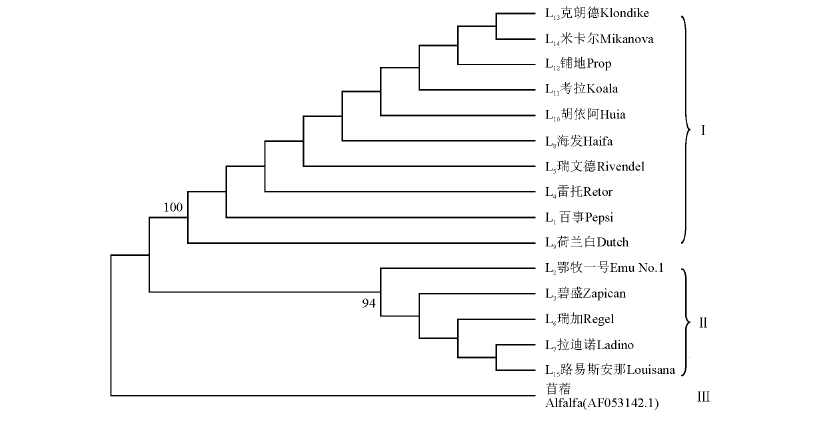 | 图3 基于ITS序列的15个白三叶品种系统发育树Fig.3 Phylogenetic tree of 15 white clover cultivars based on ITS sequences |
ISSR分子标记与ITS均将15个白三叶品种分为两大类, 且每一类包含的白三叶品种相似, 说明本研究结果可靠。而ISSR分子标记在分为两大类的基础上, 又将第Ⅰ 大类分为3个亚类, 分类更为具体。由于ISSR是一个高度可变的DNA区域, 可能产生更多的多态性条带[16], 而白三叶ITS序列变异小, 造成了其品种间最小遗传距离偏小, 因此, 可以验证ISSR遗传分析结果的可靠性, ITS更适于研究科属的区分。ISSR标记能有效揭示核桃(Juglans regia)品种间的遗传多样性, 为核桃品种资源的种质鉴定和遗传亲本的选配提供了可靠的技术支撑和科学依据[17]。刘峥和张汉尧[18]利用ITS序列分析成功构建了旋花科14个属的的系统进化树。
ISSR和ITS聚类分析表明, 选择的白三叶品种具有一定的地域性分布规律, 并与引种历史具有较强的关联性。据记载, 白三叶的种植历史悠久, 16世纪后期, 荷兰开始用种子种植。17世纪传入英国, 随后传入美国、新西兰等国[19]。来自丹麦、澳大利亚、新西兰和荷兰的白三叶品种聚在一起, 澳大利亚和新西兰都属于大洋洲, 由于地理关系聚在一起, 而后再与新西兰和丹麦聚在一起。而美国、阿根廷和中国的聚为一类, 推测是由于育种学家在育种过程中首先选择当地或者相近地区引进的白三叶种质材料进行白三叶新品种的选育, 但同时美国的瑞加和中国的鄂牧一号聚在一起, 则是由于鄂牧一号是在瑞加的基础上进行综合选育的, 而且鄂牧一号在国内推广面积不大, 变异较小, 还和原种保持着一定的亲缘关系。由于供试白三叶品种有限, 可以推测, 随着选择白三叶品种的增多, 将会有不同地区的白三叶因不同地区原种的选择而聚在一起。研究表明, 利用形态特征、SRAP、同工酶、ISSR进行遗传多样性分析能够将大部分地理区域临近的种质材料聚在一起[19, 20, 21]。
在ISSR分类中, 首先是根据地理位置将白三叶品种聚为一类, 小叶型的瑞文德和大叶型的克朗德单独与中叶型白三叶百事和米尔卡共同构成了第1亚类, 说明亲缘关系与叶形大小相关性不是很强。
本研究表明, 白三叶品种的遗传关系与地理分布具有较大的相关性, 同时也与白三叶引种历史具有较强的相关性, 尤其栽培品种间的平均相似系数相对较大[22], 在新品种选育过程中首先要考虑品种来源, 其次考虑白三叶的叶形等相关形态性状, 对目标性状进行选择育种。而同时ISSR和ITS均能有效地对白三叶品种遗传多样性进行分析, 且ISSR可以更加细致地对其进行分类, 以后可以在更多白三叶的相关研究中加以应用。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|