

{kind=link}
{kind=link}
{kind=link}
不同光环境下兴安胡枝子叶片光合和叶绿素荧光参数的光响应特点
[李鑫1  , 张会慧
, 张会慧2 , 张秀丽1 , 许楠3 , 胡举伟1 , 逄好胜1 , 滕志远1 , 孙广玉1 ]
, 张会慧]
|
|


第一作者:李鑫(1986-),女,黑龙江绥化人,在读博士生,主要从事植物生理生态学研究。E-mail:swx05256lx@126.com
为了解兴安胡枝子( Lespedeza daurica)对不同森林光环境的适应性,分别测定了林下和林缘兴安胡枝子叶片的光合气体交换参数和叶绿素荧光参数的光响应曲线。结果表明,在低光强下,林下兴安胡枝子叶片的净光合速率(Pn)和表观量子效率(AQY)明显高于林缘,即林下兴安胡枝子叶片对弱光的捕获和利用能力明显高于林缘,但林下兴安胡枝子在相对较强光强下的光合能力明显低于林缘,表现为较低的光饱和点(LSP)和净光合速率(Pmax),随着光强的增加而减少,林下兴安胡枝子叶片的气孔导度(Gs)和蒸腾速率(Tr )增加幅度也明显低于林缘,但林下兴安胡枝子具有相对较高的水分利用效率。低光强下,林下和林缘兴安胡枝子叶片的实际光化学效率(ФPSⅡ)和电子传递速率(ETR)均无明显差异,但当光强超过400 μmol·m-2·s-1后,林缘兴安胡枝子叶片的ФPSⅡ和ETR均明显高于林下,但是林下兴安胡枝子叶片的热耗散能力明显高于林缘。兴安胡枝子叶片的光合特性对不同光环境具有较强的适应能力。
, Zhang Hui-hui
In order to understand the adaptability of Lespedeza davurica on different forest light environmental, photosynthetic gas exchange and light response curve of chlorophyll fluorescence parameters were detected respectively. The results showed that Pn and AQY of L. davurica leaf growth in understory were significantly higher than that of the forest edge under low light intensity, which indicated that the ability of capture and utilization of weak light growth in understory were significantly higher than that in the edge of forest. On the contrary, the ability of capture and utilization of strong light growth in understory was significantly lower than that in the edge of forest, in which the value of LSP and Pmax were lower. The increased extent of GS and Tr of L. davurica leaves understory were lower than the edge of the forest with increasing the light intensity, but L. davurica grown in forest has relatively higher water use efficiency. Under low light intensity, the parameters of ФPSⅡ and ETR were both not different between understory and forest edge. But the parameters of ФPSⅡ and ETR for those grown in forest edge were higher than understory when the intensity was more than 400 μmol·m-2·s-1, ETR and LCP of L. davurica leaves were significantly higher than those understory. The heat dissipation capability of understory was higher than those grown in forest edge. In conclusion, the results of this study showed that the ability of L. davurica for adapting different light environments were strong.
光是植物进行光合作用以及正常生长的前提, 也是影响植物生长、形态建成、存活和分布的重要生态因子[1]。植物叶片光合色素的含量和比例、光能吸收和利用能力等均明显受光照条件的影响, 生长在弱光下的植物常具有较高的比叶面积和叶绿素含量以吸收较多的光能, 维持植物光合能量供应[2]。但是生长在过高光强下的植物, 特别是具有阴生特点的植物, 光胁迫又是限制其光合作用的重要因素之一。研究发现, 植物处于光胁迫下光合作用光抑制造成约10%的碳损失[3, 4]。因此, 光对植物的光合作用及生长是一把双刃剑, 植物具有对不同光环境特别是变化光环境的适应能力, 是在长期物种进化和自然生境选择中不被淘汰的关键。森林是一个特殊的生态环境, 由于林缘和林窗的存在, 不同区域的光、温、湿等生态因子存在明显的异质性[5], 虽然温度和湿度等生态因子对植物存在潜在的影响, 但不同生境中光照强度的差别最大, 往往相差数倍甚至数十倍, 为影响植物生长及存活的主效应因子[6]。随着砍伐和林火等人为因素的干扰, 连片的森林遭到破坏, 导致林缘的增加, 林缘效应对森林的影响程度逐渐加大。林缘效应使植物群落更新演替加快, 生长在林下的阴生植物逐渐退化, 被喜光的阳生或半阴生先锋植物替代[7]。植物具有适应这种光环境变化的适应能力是其能够得以保存的关键。
兴安胡枝子(Lespedeza daurica)为豆科(Leguminosae)蝶形花亚科(Papilionatae)胡枝子属的半灌木或灌木, 胡枝子具有较强的抗旱、耐寒、耐瘠薄和耐酸等特点, 可以作为优良的水土保持和绿化美化树种[8, 9], 胡枝子根系发达, 且含根瘤菌, 在北方干旱半干旱地区植被恢复中具有重要的作用[10]。另外, 胡枝子萌芽力强、具有较好的耐刈割和适口性特点, 叶片中粗蛋白质和粗脂肪含量较高, 其嫩枝可以作为优质青饲料, 具有较高的经济价值[11]。在大兴安岭地区, 胡枝子常常是优势种蒙古栎(Quercus mongolica)林的伴生种, 具有明显的耐阴性。本研究在自然环境下研究林下和林缘兴安胡枝子叶片的光合气体交换和叶绿素荧光参数的光响应特点, 探讨兴安胡枝子光合特性对光环境的适应机制, 以期了解兴安胡枝子在群落演替中的适应机制和策略, 为兴安胡枝子的种质保存和群落优化配制提供基础数据。
试验于2013年7月在位于大兴安岭林区南缘的加格达奇林区进行。试验地点位于大兴安岭伊勒呼里山南部东坡(124° 01'-124° 07' E, 50° 15'-50° 18' N), 地带性土壤为暗棕壤。该地区属于寒温带大陆性季风气候[12]。试验地选取蒙古栎林为主的乔木次生林, 郁闭度较高, 林下伴生有兴安胡枝子、苔草(Carex tristachya)、地榆(Sanguisorba officinalis)、芍药(Paeonia lactiflora)、苦参(Sophora flavescens)、藜芦(Veratrum linn)、单花鸢尾(Iris uniflora)、野豌豆(Vicia sepium)等相对耐阴植物。林缘与林下相似, 除主要分布兴安胡枝子、苔草、苦参等植物外, 还有大量山杏(Prunus armeniaca)、裂叶蒿(Artemisia tanacetifolia)、兔儿伞(Syneilesis aconitifolia)、委陵菜(Potentilla aiscolor)、唐松草(Thalictrum aquilegifolium)、败酱(Patrinia rupestris)、矮蒿(Artemisia lancea)、蚊子草(Filipendula palmata)、狼毒大戟(Euphorbia fischeriana)、山莴苣(Mulgedium sibiricum)和黄岑(Scutellaria baicalensis)等中生或相对喜光的植物。
光响应曲线的测定:利用Li-6400光合测定系统, 将仪器自带光源分别设定光强(PFD)为2 000、1 800、1 600、1 400、1 200、1 000、800、600、500、400、300、250、200、150、100、75、50、25和0 μ mol· m-2· s-1, 按照光强由高到低的顺序分别测定林缘和林下胡枝子完全展开功能叶片的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci), 并计算水分利用效率(WUE), WUE=Pn/Tr, 绘制Pn-PFD、Gs-PFD、Tr-PFD、Ci-PFD和WUE-PFD响应曲线, 用Photosynthesis Work Bench程序软件求得Pn-PFD响应曲线的拟合系数(R2)以及表观量子效率(AQY)、暗呼吸速率(Rd)、光补偿点(LCP)、光饱和点(LSP)、光饱和时的净光合速率(Pmax)。
叶绿素荧光曲线的测定:用暗适应夹对不同光环境下树冠外围兴安胡枝子完全展开功能叶片进行暗适应30 min后, 利用便携调制式荧光仪(FMS-2, Hansatech, 英国)测定最大荧光(Fm)、初始荧光(Fo), 然后用FMS-2的内置光源给叶片施加不同的作用光, 光照强度(PFD)分别设为100、200、400、600、800、1 000和1 200 μ mol· m-2· s-1, 每个光强下适应 3 min后测定光适应下的叶片的荧光参数, 即, 光适应下的最小荧光(Fo')、光适应下的最大荧光(Fm')及稳态荧光(Fs), 分别计算实际光化学效率(Ф PSⅡ )、电子传递速率(ETr)、光化学淬灭系数(qP)和非光化学淬灭系数(NPQ), 其公式分别为Ф PSⅡ =(Fm/Fs)/Fm', ETr=0.5× 0.85× Ф PSⅡ × PFD, qP=(Fm'-Fs)/(Fm'-Fo'), NPQ=(Fm/Fm')/Fm'[13]。
运用Excel 2003和SPSS 7.05软件进行数据统计分析, 采用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)比较不同数据组间的差异。
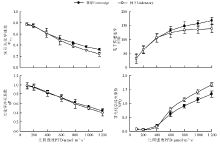
不同光环境下兴安胡枝子叶片的Pn-PFD曲线明显不同, 其中在光强低于500 μ mol· m-2· s-1时, 林下兴安胡枝子叶片的Pn明显高于林缘, 即林下兴安胡枝子叶片的光合能力对弱光的适应性较强。随着光强的增加, 林缘兴安胡枝子叶片的Pn增加幅度明显大于林下, 并且随着光强的增加, 林下兴安胡枝子在接近1 000 μ mol· m-2· s-1时基本达到光饱合, 但此时林缘兴安胡枝子Pn随着光强的增加仍呈明显的增加趋势(图1)。利用Photosynthesis Work Bench程序软件分析林缘和林下兴安胡枝子叶片的Pn-PFD响应曲线的拟合系数分别为0.99和1.00, 均可以较好拟合曲线的性质。林下兴安胡枝子叶片的AQY高于林缘57.14%(P< 0.05), 但其Rd、LCP、LSP和Pmax则分别低于林缘22.78%(P< 0.01)、49.46%(P< 0.01)、21.73%(P< 0.05)和32.33%(P< 0.01)(表1)。
 | 图1 不同光环境下兴安胡枝子叶片的净光合速率光响应曲线Fig.1 Response of net photosynthetic rate to illumination intensity in leaves of L. daurica under different light environments |
| 表1 不同光环境下兴安胡枝子叶片的光合参数 Table 1 Photosynthetic parameters in leaves of L. daurica under different light environment |
不同光环境下兴安胡枝子叶片的Gs和Tr的变化趋势相似。在弱光下, 林缘和林下兴安胡枝子叶片的Gs和Tr无明显差异, 但随着光强的增加, 林缘兴安胡枝子叶片的Gs和Tr增加幅度明显高于林下, 并且二者差异幅度也随着光强的增加而变大(图2)。
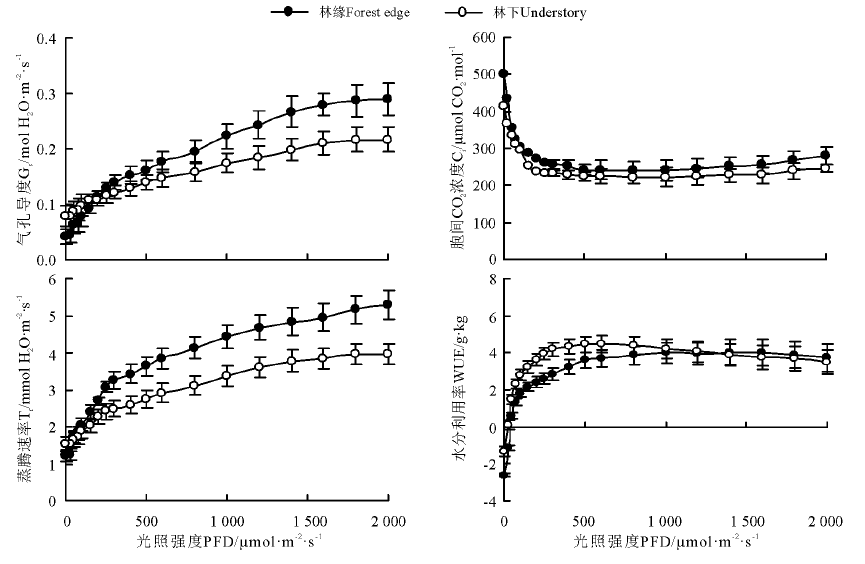 | 图2 不同光环境下兴安胡枝子叶片的气孔导度、蒸腾速率、胞间CO2浓度和水分利用效率光响应曲线Fig.2 Response of stomatal conductance, transpiration rate, intercellular CO2 concentration and water use efficiency to light inlensity in leaves of L. daurica under different light environments |
在弱光下, Ci随着光强的增加呈降低趋势, 之后Ci变化较为平缓, 不同光强下林缘和林下兴安胡枝子叶片的Ci均表现为林缘大于林下, 但差异较小。在光强小于500 μ mol· m-2· s-1时, 林下兴安胡枝子叶片的WUE明显高于林缘, 但随着光强的增加二者差异逐渐变小, 并且当光强达到1 000 μ mol· m-2· s-1后二者基本相同(图2)。
随着光强的增加, 兴安胡枝子叶片的Ф PSⅡ 和qP呈降低趋势, 而ETR和NPQ则逐渐增加, 并且在光强低于400 μ mol· m-2· s-1时, 林下和林缘兴安胡枝子叶片的各叶绿素荧光参数均无明显差异, 当光强超过400 μ mol· m-2· s-1时, 随着光强的增加, 林下兴安胡枝子叶片的Ф PSⅡ 和ETR逐渐低于林缘, 并且随着光强的增加, 差异逐渐变大。不同光强下林下和林缘兴安胡枝子叶片的qP差异程度较小, 仅表现为高光强下林缘稍高于林下。在低于400 μ mol· m-2· s-1光强下, 林下和林边缘兴安胡枝子叶片的NPQ均维持在较低水平, 并且二者也无明显差异, 但光强超过400 μ mol· m-2· s-1后, 两种光环境下叶片的NPQ均迅速增加, 且林下叶片的NPQ明显大于林缘(图3)。
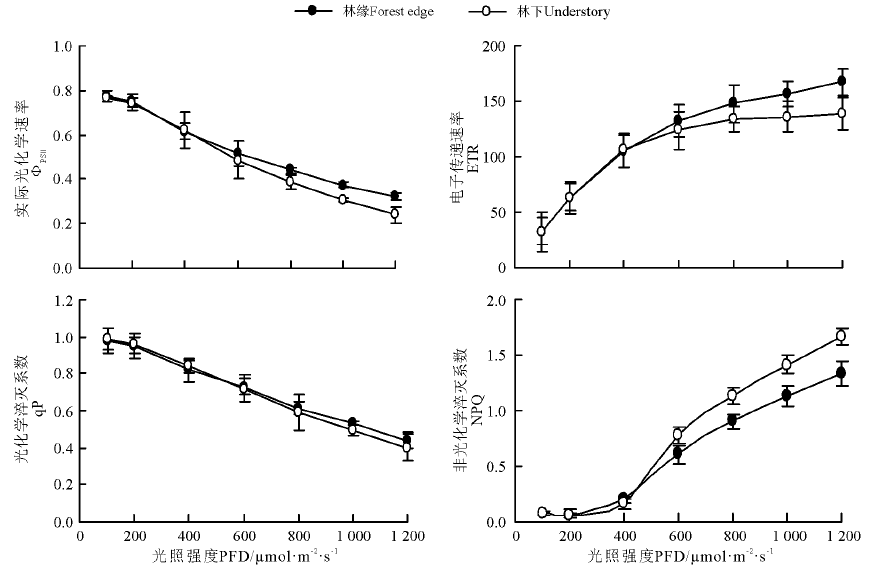 | 图3 不同光环境下兴安胡枝子叶片的叶绿素荧光参数光响应曲线Fig.3 The chlorophyll fluorescence parameters response to illumination intensity in leaves of L. daurica under different light environments |
光合作用是植物获取物质和能量的基础, 并且光合作用也是对环境极为敏感的生理过程之一。在不同光环境下, 植物叶片会通过改变比叶面积、改变色素的含量和比例来调整光能的吸收和利用, 以维持正常光合的能量所需以及防止过剩光能的过量积累[14], 弱光环境下常常因同化力的短缺而限制光合碳同化, 表现为净光合速率的降低[15]。本研究中, 林下兴安胡枝子叶片的Pn在作用光强低于500 μ mol· m-2· s-1时显高于林缘, 并且其AQY也明显高于林缘, AQY是植物对光能利用效率的重要指标, 反映了植物捕获光量子特别是弱光下捕获光量子用于光合作用的能力[16], 即林下兴安胡枝子叶片对弱光的利用能力明显高于林缘, 这是植物长期生长在相对弱光下的一种适应能力, 这可能与林下兴安胡枝子叶片的叶绿素含量较高, 对弱光的捕获能力增强有关, 或较高的叶绿素含量导致光的反射和透射损失较少有关, 也可能是由于林缘兴安胡枝子叶片受到了强光的伤害, 这与曾伟等[17]的研究结果相似。但由于林下兴安胡枝子叶片长期生长在弱光下, 光合作用的关键酶和气孔等具有适应弱光环境的能力, 因此导致其在光强相对较强时光合能力明显低于林缘, 表现为LSP和Pmax降低, Pmax体现了植物潜在的光合能力, 而LSP反映了植物对高光强的利用能力[18], 因此, 本研究表明, 生长在林缘的兴安胡枝子与林下不同, 对高光强的利用能力明显增强。另外, 林下兴安胡枝子叶片的Rd明显低于林缘, 这有利于弱光下兴安胡枝子的净光合积累, 是一种对弱光环境的适应机制, 这与齐欣等[19]在蒲桃属(Syzygium)植物适应弱光上的研究结果相似。
随着光强的增加, 林下和林缘兴安胡枝子叶片的气孔逐渐张开, 导致其蒸腾速率也随着气孔导度的增加而增加, 二者呈明显的正相关关系, 即蒸腾速率主要受气孔因素的影响。但是虽然随着光强的增加兴安胡枝子叶片的气孔导度增加, 但在低于500 μ mol· m-2· s-1下Ci随着光强的增加却呈降低趋势, 之后变化较为平缓, 也没有随着气孔导度的增加而增加, 说明随着光强的增加, 兴安胡枝子叶片光合暗反应逐渐活化, 净光合速率Pn增加, 对CO2的利用能力增强, 从而导致细胞间的CO2浓度降低。另外, 弱光下兴安胡枝子叶片的WUE呈增加趋势, 这与随着光强的增加, 兴安胡枝子碳同化的速度大于水分蒸腾速率有关。不同光环境下比较, 在低于200 μ mol· m-2· s-1光强下, 林下光安胡枝子叶片的Gs和Tr均稍高于林缘, 这与其长期生长在弱光环境下有关, 但是随着光强的增加, 林缘兴安胡枝子叶片的Gs和Tr增加幅度明显大于林下, 即林缘光环境下兴安胡枝子对相对强光的利用能力大于林下。并且在低于1 000 μ mol· m-2· s-1光强下林下兴安胡枝子叶片的WUE明显大于林缘, 这除了与弱光下(约低于500 μ mol· m-2· s-1)林下兴安胡枝子碳同化能力较强有关外, 还与其在500~1 000 μ mol· m-2· s-1光强下林下叶片的蒸腾速率较低有关。
叶绿素荧光技术是反映植物光合作用内在机理的重要手段之一。本研究中, 随着光强的增加, 兴安胡枝子叶片的Ф PSⅡ 呈降低趋势, 而ETR则呈增加趋势, Ф PSⅡ 的降低原因可能与随着光强的增加兴安胡枝子叶片PSⅡ 反应中心逐渐关闭或激发能以其它途径耗散有关[20, 21]。100~400 μ mol· m-2· s-1的光强下, 林下和林缘兴安胡枝子叶片的Ф PSⅡ 和ETR均无明显差异, 即在弱光作用下林下和林缘兴安胡枝子叶片吸收光能用于光合电子传递的比例无明显差异, 但此时林下兴安胡枝子叶片的碳同化能力却明显高于林缘, 造成此现象的原因可能与光合电子流分配的改变有关, 即弱光下虽然林下兴安胡枝子叶片的线性电子流与林缘相同, 但线性电子流中除了光呼吸、氨同化和环式电子流等之后的组分才与净光合速率具有明显相关性, 而在弱光下林缘兴安胡枝子可能除了将吸收的光能用于光化学反应外, 可能还启动了其它耗能途径, 从而造成其净光合速率明显低于林下。但是当光强超过400 μ mol· m-2· s-1后, 林缘兴安胡枝子叶片的Ф PSⅡ 和ETR均明显高于林下, 并且通过ETR-PFD的光响应曲线也可以看出, 林缘兴安胡枝子叶片的ETR光饱和点以及光饱和时的最大电子传递速率也明显高于林下, 即林缘兴安胡枝子叶片对强光的利用能力明显高于林下, 这与净光合速率的变化结果相符。qP在一定程度上反映了PSⅡ 反应中心的开放程度, 而NPQ则与依赖叶黄素循环的热耗散呈正相关关系[22, 23], 本研究中, 随着光强的增加, 兴安胡枝子叶片的qP呈降低趋势, 而NPQ则逐渐增加, 即PSⅡ 反应中心随光强的增加逐渐关闭, 但在反应中心关闭, 而在过剩激发能存在的情况下, 兴安胡枝子叶片主要通过热耗散来保护PSⅡ 的正常生理功能[24]。虽然在相对高光强下林下兴安胡枝子叶片的Pn、Ф PSⅡ 和ETR均明显低于林缘, 但其qP差异却较小, 说明在强光下造成林下兴安胡枝子叶片光合能力低于林缘的原因并不是强光导致PSⅡ 反应中心关闭程度不同, 可能与其光能的利用方式不同有关。通过NPQ的光响应曲线也可以看出, 随着光强的增加, 即弱光下的兴安胡枝子在高光下主要通过热耗散来利用过剩光能, 以保护PSⅡ 反应中心活性。林下兴安胡枝子叶片的热耗散能力明显高于林缘, 这与生长在相对弱光下林下兴安胡枝子叶片对高光强的利用能力较低有关。
兴安胡枝子对不同光环境具较强的适应能力, 在林下, 兴安胡枝子对弱光的利用能力较强, 并且具有相对较高的水分利用效率, 林缘兴安胡枝子在强光下不但具有相对较高的碳同化能力, 而且叶片的PSⅡ 反应中心光化学活性也明显高于林下。兴安胡枝子光合能力对不同光环境的适应性是导致其在大兴安岭地区分布较广的重要原因之一。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|