

{kind=link}
{kind=link}
{kind=link}
白三叶返园对苹果园土壤微生物群落的影响
[魏倩倩1  , 杨文权
, 杨文权1 , 韩明玉2 , 慕小倩1 , 寇建村3 ]
, 杨文权, 寇建村]
|
|


第一作者:魏倩倩(1992-),女,陕西宝鸡人,在读硕士生,主要进行土壤微生物多样性研究.E-mail:weiqianqian1641@126.com
为了解苹果( Malus pumila)园中覆盖和埋置白三叶( Trifolium repens)对土壤微生物群落的影响,采用尼龙袋法将新鲜的白三叶覆盖和埋置到苹果园中,分别在白三叶降解1,3,6个月时,利用Biolog微平板法检测土壤微生物群落的功能多样性.结果表明,覆盖和埋置白三叶均可使苹果园土壤微生物代谢活性显著提高( P<0.05),微生物的多样性除6个月外显著增加,丰富度显著增加,均匀度显著降低( P<0.05),群落结构发生变化,且覆草处理大于埋草,同时,白三叶降解时间的长短也影响土壤微生物群落.土壤微生物群落主成分分析结果显示,不同返园方式间主要对羧酸类碳源的利用有差异,而导致不同降解时间土壤微生物群落代谢差异的主要是糖类.说明在苹果园覆盖和埋置白三叶可改变土壤微生物群落,而覆盖白三叶对微生物群落结构的改善情况显著优于埋置白三叶.
In order to understand the effects of burying and covering white clover ( Trifolium repens) in apple ( Malus pumila) orchard on the soil microbial community, the soil microbial community functional diversity was analyzed by Biolog microplate method after 1, 3 and 6 months treatments which fresh white clover was buried or covered in apple orchard by litter nylon bag, respectively. The results indicated that both covering and burying white clover could significantly elevate ( P<0.05) the soil microbial metabolism activities, the microbial Shannon-Winner diversity index(except 6 months) and richness index while significantly reduced ( P<0.05)uniformity index and changed soil microbial community structure. The effects of covering white clover were greater than that of burying. And the time of white clover degradation also had effects on soil microbial community. Principal component analysis of soil microbial community showed that the major differences between covering and burying were the utilization of carboxylic acids, whereas the major differences of microbial metabolism activities between degradation periods were sugars. These results indicated that both covering and burying white clover can change the soil microbial community in apple orchards, and the effects of covering were greater than that of burying.
黄土高原是我国苹果(Malus pumila)的主产区, 但是长期以化肥为主的施肥模式导致土壤肥力退化, 养分失调以及有机质含量低等一系列问题, 严重制约了我国苹果产业的发展[1, 2].近年来, 果园生草模式逐渐发展成为黄土高原苹果园的一种新的土壤管理模式, 可以有效改善苹果园土壤养分状况[3, 4].同时, 将果园所生牧草适时刈割后返园, 可将其光合作用固定的物质以及从土壤中吸收的养分再返还土壤, 使物质能量得以循环利用[1].但是, 目前对牧草的返园方式以及牧草降解对土壤影响方面的研究还鲜有报道[5].
土壤微生物群落的组成和代谢状态是土壤质量变化最敏感, 最有潜力的生物学指标[6].相关研究表明, 植物残体降解对土壤微生物群落结构有重要影响[7, 8, 9], 植物残体分解与土壤微生物群落结构功能变化是相互影响, 相互制约的过程[7].Biolog微平板法是测定土壤微生物功能多样性常用的, 简单快速的方法, 该方法是通过测定微生物对单一碳源的代谢利用情况, 来描述土壤微生物群落结构的差异和功能多样性[10, 11, 12].为掌握果园牧草不同的返园方式对土壤微生物群落多样性的影响, 本研究以黄土高原苹果园最适宜的草种白三叶(Trifolium repens)为材料[2, 13], 刈割后以覆盖和埋置的方法返园, 通过Biolog微平板法, 研究白三叶不同的返园方式对土壤中微生物群落功能多样性的影响, 以期为果园牧草的合理利用提供科学的理论依据.
试验地位于陕西省乾县梁山乡双矮苹果园, 地处关中平原中段北侧, 渭北高原南缘, 108° 20' E, 34° 62' N, 海拔830 m, 属温带大陆性季风气候.年均气温13.1 ℃ , 年降水量573~590 mm, 无霜期244 d.春季气温变化大, 雨水不均.夏季炎热, 降水量大且分布不均.秋季气温温和多雨.冬季寒冷干燥.2006年春季建园, 双矮栽培, 种植的苹果树种为礼泉短富, 行距2.5 m, 株距1 m, 果园管理水平成熟.土壤基础差, 养分含量低, 熟土层薄[14].
2014年7月26日刈割苹果园中种植的白三叶, 装入尼龙袋中, 每袋装鲜草200 g.7月27日, 在苹果树行的中间位置, 每隔一定距离, 挖深40 cm, 宽30 cm的沟, 将沟内取出的土壤混匀, 备用.试验设覆草和埋草两个处理.覆草处理:将混匀的土装入高40 cm, 长, 宽均为30 cm的软塑料花盆内, 放入沟内, 使其与地面保持平整, 将装有白三叶的尼龙袋压平覆盖在容器土壤表面; 埋草处理:先在容器内装入15~20 cm深的土, 再把尼龙袋放入容器中, 然后, 用土把容器装满, 容器表面与周围地面保持平整.对照:不放白三叶的土壤装满容器.每处理重复5次.
分别于2014年8月27日, 10月27日, 2015年1月27日进行土壤样品采集.土壤样品采集时, 混匀容器内的土壤, 除去可见植物残体及土壤动物, 用四分法留取土样后一部分装入铝盒中进行土壤含水量测定, 再取一部分用无菌自封袋封装, 放入装有冰袋的保温箱, 运回实验室, 进行简单的过筛处理后混匀, 密封, 置于4 ℃ 冰箱中保存, 用于土壤微生物群落功能多样性的测定.
土壤微生物功能多样性利用Biolog生态测试板(EcoPlatesTM, 美国 Matrix Technologies Corporation)进行测定, 方法参考Garland等[10]的方法.平均颜色变化率(Average Well Color Development, AWCD)的计算公式为AWCD=∑ (Ci-R)/3, 式中, Ci为每个孔在590 nm下的吸光值, R为A1孔在590 nm下的吸光值.如果Ci-R< 0, 则在计算中记为0.一个ECO微孔板上的碳源有3个重复, 则AWCD值为3次重复的平均值.本研究取培养72 h的AWCD值进行微生物功能多样性分析, 其中包括Shannon-Winner指数(H), H=-∑ Pi(lnPi), 式中, Pi表示第i孔的吸光值与对照孔的吸光值之差与整个微孔板相对吸光值的比值, Pi=(Ci-R)/∑ (Ci-R); 丰富度指数(S)为被利用的碳源的数目, AWCD≥ 0.2时代表该孔中的碳源被利用; 均匀度指数(E)的计算公式为E=H/lnS.
使用Microsoft Excel 2013对数据进行初步处理, 使用Statgraphics软件对数据进行正态分布和方差齐次性检验, 通过检验后对数据进行方差分析(ANOVA), 当ANOVA的F检验的P值小于0.05时认为均值间的差异显著, 然后利用OriginPro 8.5作图.主成分分析(Principal Component Analysis, PCA)在R软件中利用prcomp()命令完成.
平均颜色变化率(AWCD值)反映了土壤微生物在培养过程中的代谢活性大小.AWCD值越高说明微生物群落数量越庞大, 代谢活动越强[15].因此, 考察不同处理不同时间的土壤微生物在培养过程中的AWCD值变化, 可以明确不同处理不同时间对土壤微生物群落代谢活动的影响.在培养的240 h内, AWCD值总体呈现不断上升的趋势, 前120 h快速增长, 120 h后增长速度逐渐放缓, 存在明显的对数期和平台期(图1).覆草处理1个月时的AWCD值一直高于埋草处理1个月时的, 覆草和埋草均高于对照.3和6个月的规律与此相同.比较同一处理3个不同降解时间的AWCD值可以发现, 6个月要明显低于1和3个月.由此可以得出, 覆草处理和埋草处理均可以提高土壤微生物群落数量和代谢活性, 并且覆草处理高于埋草处理, 并且在1和3个月时这种趋势明显, 6个月时减弱.
 | 图1 土壤微生物群落AWCD随着培养时间的变化Fig.1 AWCD changes with different incubation time |
物种多样性是物种丰富度和物种均匀度的综合指标, 可以反映微生物群落结构和功能状态, 是表征生态系统是否健康稳定的重要标志[16].Shannon-Winner指数即多样性指数, 其值越高说明微生物群落功能多样性越高[17].丰富度就是群落中物种数目的多少.均匀度反映的是群落中全部物种的个体数目的分配情况.取培养72 h的土壤微生物群落AWCD值作多样性分析, 分析了Shannon-Winner多样性指数, 丰富度指数, 均匀度指数(表1), 结果表明, 处理及处理时间对Shannon-Winner多样性指数, 丰富度指数, 均匀度指数和72 h AWCD值均有影响.处理1个月时, 覆草的72 h AWCD值, 土壤微生物群落的Shannon-Winner多样性指数和丰富度指数显著高于埋草和对照(P< 0.05), 并且72 h AWCD值和丰富度指数埋草显著高于对照, 3个月和6个月时的规律与此基本相同.而均匀度表现为覆草和埋草显著小于对照, 覆草小于埋草.这说明了覆草和埋草可以提高土壤微生物群落的多样性和丰富度, 降低均匀度, 但是埋草效果不如覆草.
| 表1 土壤微生物群落多样性指数和72 h AWCD Table 1 Diversity indices of soil microbial communities and AWCD of 72 h |
分析同一处理不同降解时间各个指数的大小情况可以发现, 对照在3个时间点的72 h AWCD值, Shannon-Winner指数和丰富度指数均没有显著差异(P> 0.05), 3个月的均匀度指数显著低于1和6个月.覆草后3个时间点的Shannon-Winner指数和均匀度指数没有差异(P> 0.05), 而丰富度指数6个月的显著(P< 0.05)低于1和3个月的.埋草Shannon-Winner指数1和3个月没有显著差异(P> 0.05), 但是均显著(P< 0.05)高于6个月的, 丰富度指数在3个时间点间的差异显著(P< 0.05), 3个月> 1个月> 6个月, 均匀度指数在3个月时最低, 6个月时最高.
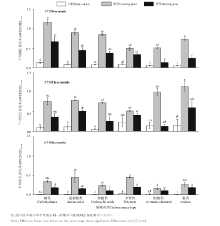
不同的土壤微生物群落对不同类型的碳源利用有差异.ECO板上共有31种碳源, 分为糖类, 氨基酸类, 羧酸类, 多聚物类, 酚酸类, 胺类共六大类.在各个时间, 覆草对各类碳源的代谢水平都要高于埋草, 且基本都差异显著(P< 0.05), 并且所利用的主要碳源类型有所不同(图2).1个月时, 覆草和埋草均以糖类为主要碳源利用类型, 其次是氨基酸类和羧酸类; 此时, 对照利用的主要碳源为糖类和多聚物.3个月时, 对照利用最多的是多聚物和胺类, 而覆草主要碳源利用类型变成了胺类和酚酸类, 埋草则主要利用了胺类和氨基酸类.6个月时覆草, 埋草和对照都主要利用的是多聚物, 但是覆草还主要利用了氨基酸类, 埋草对各类碳源均有所利用, 差异不明显.
 | 图2 土壤微生物六大类碳源代谢分析Fig.2 Six carbon source types utility analysis of soil microbial community |
上述结果分别分析了不同处理不同降解时间土壤微生物群落在利用碳源总体和不同碳源类型两个水平上的差异, 而主成分分析(PCA)可以反映微生物利用碳源的更详细情况.各个样本分布在了PCA图上不同的位置, 表明各样本利用碳源的能力有所不同, 而这种差异与各样本在利用聚集在PC1轴和PC2轴上的碳源有关.取培养72 h的土壤微生物群落AWCD值作主成分分析(图3), 31个成分因子中前6个的累积方差贡献率达到82.61%, 从中提取前两个主成分PC1, PC2, 它们的累积方差贡献率达到60%, 可以解释60%的单一碳源变量的数据变异.3个降解时间的对照都聚集在了一起, 1个月的覆草和埋草位于第3象限, 3个月的埋草和覆草位于第2象限, 6个月的埋草和覆草位于第1象限.PC1轴可以将3个时间点的对照, 覆草和埋草分开, 解释了49%的总变异, 代表了由不同处理引起的变异.PC2轴可以将覆草的3个不同时间点分开, 将埋草的1与3, 6个月分开, 而无法将对照的3个时间点区分开.通过比较不同处理点之间的距离可知(图3), 1和3个月不同处理之间对31种底物利用的差异大于6个月的, 6个月时不同处理对底物的利用趋于一致, 同时覆草3个时间点对31种底物的利用差异大于埋草.根据PCA因子载荷得分表可知(表2), 对PC1贡献大的10种碳源中4种碳源属于羧酸类, 糖类和氨基酸类各占两种.对PC2贡献大的碳源也有10种, 其中5种属于糖类, 两种属于胺类.
 | 图3 土壤微生物群落碳源利用主成分分析Fig.3 PCA of carbon utilization of the soil microbial community |
| 表2 与PC1和PC2相关性高的主要碳源 Table 2 Substrates with high correlation coefficients for PC1 and PC2 in PCA |
白三叶作为外源有机物质施入土壤后, 可以被土壤微生物利用, 增强土壤微生物的活动, 反过来, 这些微生物又可以加速白三叶的分解, 促进营养元素的释放以及营养元素在土壤生态系统中的循环[18, 19].白三叶刈割后新鲜的植物残体在降解过程中, 土壤微生物不仅是降解的主要执行者, 而且还受降解过程的影响, 因此, 随着分解过程的进行, 土壤微生物的种类和数量以至于功能多样性都在不断变化中, 水分, 温度, 光照等生态因素也会影响果园土壤微生物群落[20, 21].降解前期主要是糖类, 蛋白质, 有机酸, 苯酚等可溶性有机组分以及K, Ca, Mg, Mn等无机组分的快速损失, 后期逐渐开始分解较难分解的大分子物质, 如纤维素, 木质素等[22, 23].本研究对比了覆盖和埋置两种白三叶的返园方式, 结果显示, 覆草和埋草均可以提高土壤微生物代谢活性, Nair和Ngouajio[24]的研究也表明, 土壤中施入有机物质可以加速土壤微生物的代谢.但是覆草提高的程度要高于埋草, 可能是由于覆草的白三叶直接暴露在自然环境中, 阳光, 雨水等自然因素能够加速其分解, 同时充足的氧气能够促进降解相关的好氧微生物的快速繁殖.与埋草相比, 覆草提高了土壤微生物的多样性和丰富度, 降低了土壤微生物均匀度, 表明覆草处理更利于某些参与白三叶降解的菌株成为优势菌株.
除此之外, 降解时间对微生物群落结构的影响也非常明显.随着降解时间的增加, 1和3个月的微生物代谢活性没有显著的差异, 但是均大于6个月, 该结果与郭剑芬等[23]认为的植物残体前期分解速率较快, 后期分解速率较慢的观点吻合, 也可能与6个月取样时已经进入冬季, 气候寒冷使微生物活动减弱.有研究表明, 随着气温降低, 植物残体的分解速率呈指数降低趋势[25].同时, 温度可以影响部分微生物的活性, 进而使植物残体中的特定组分降解速度发生改变[26].尽管覆草和埋草主要利用的碳源随着降解时间的推移发生改变, 但是对照处理主要利用的碳源并没有改变, 说明温度对碳源利用的影响在本研究中影响较少.PCA分析发现, 6个月时覆草, 埋草和对照之间的差异相比1和3个月进一步缩小, 同时6个月的覆草和埋草处理与3次对照在碳源利用上更为接近, 表明低温虽然降低了微生物代谢活性, 但是造成6个月时处理间的差异逐渐减少的主要诱因是白三叶分解后期可利用的底物逐渐减少, 覆草和埋草所处的微环境接近对照.覆草处理后, 微生物多样性指标在3个时间点没有显著变化, 而埋草处理后随着时间变化出现较大波动, 表明与埋草处理相比, 覆草处理微生物群落结构更加趋于稳定, 在较长时间内能够保持优势菌株的优势度.
此外, 通过进一步研究微生物对六大类碳源代谢的情况发现, 覆草和埋草可以改变土壤微生物主要碳源利用类型, 并且不同降解时间利用的主要碳源不同.对照在3个时间主要都以利用多聚物为主, 而覆草利用的主要碳源在逐渐变化, 从1个月时的糖类到3个月时的胺类, 再到6个月时的多聚物, 埋草处理的主要碳源利用类型和覆草处理类似.这说明白三叶的降解可以在一定程度上改变土壤微生物结构类群, 在前期由于白三叶中糖类, 胺类等营养物质的快速损失, 而增加了能够利用糖类, 胺类微生物的类群, 在6个月时可能是由于三叶草中的能快速被利用的营养物质消耗殆尽有关, 覆草和埋草的土壤微生物利用各个碳源相比对照已经没有了明显偏好.PCA分析表明, 对PC1和PC2起主要分异作用的主要碳源分别是羧酸类和糖类, 不同处理间主要对羧酸类碳源的利用有差异, 而导致不同三叶草降解时间土壤微生物群落代谢差异的主要是糖类.羧酸类物质是造成覆草, 埋草和对照之间微生物群落差异的主要因素, 与之对应的是3个不同时期3个处理之间对羧酸的利用都差异显著, 同时覆草处理对羧酸的利用都大于埋草处理, 表明覆草所处的环境更有利于降解过程中羧酸类物质的释放.降解过程中, 一部分可溶性糖在降解的早期迅速释放, 使利用其的微生物群落优势度迅速提升, 但是更多的糖类是植物结构类物质(例如纤维素等), 这些糖类物质降解较慢, 而且主要在降解后期开始降解[22, 23].因此, 糖类在降解所经历的时间较其它物质更长, 所以在不同降解时间的微生物群落结构功能中起主要分异.
总之, 覆草提高了微生物群落代谢活性以及多样性和丰富度, 使土壤微生态系统更加稳定, 使土壤养分向有利于苹果树生长的水平靠近.而且白三叶刈割后直接覆盖比埋草更省时省力利于操作.白三叶不同的返园方式对果树生长以及对苹果品质性状的影响有待后续进一步研究.
1)覆草和埋草都可以明显提高土壤微生物群落的代谢活性以及微生物群落的多样性和丰富度, 降低均匀度, 但是覆草处理的这些趋势要明显高于埋草.
2)覆草和埋草可以改变土壤微生物主要碳源利用类型, 并且不同时间利用的主要碳源不同, 表明草随着时间的降解可以不断改变土壤微生物群落结构.
3)不同处理间主要对羧酸类碳源的利用有差异, 而导致不同时间土壤微生物群落代谢差异的主要是糖类.
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|