




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
普陀山苔草植物络合素合成酶CpPCS基因克隆及其在叶中的表达分析
[谭家郎1 , 胡永霖1 , 王成龙2 , 杨占彪1  , 朱雪梅
, 朱雪梅1 ]
, 朱雪梅|
|


第一作者:谭家郎(1991-),男,湖南涟源人,在读硕士生,主要从事污染生态学研究。E-mail:tanjia1991@qq.com
以超富集植物普陀山苔草(Carex putuoshan)为材料,克隆其植物络合素合酶基因CpPCS的cDNA全长序列。同时,对其进行重金属胁迫处理,研究该基因在叶中的表达特点。结果表明,该基因cDNA序列全长1 461 bp,可编码486个氨基酸;与其它植物同源基因对比显示,它们的氨基酸序列相似性在63%左右;通过构建进化树发现,普陀山苔草CpPCS与小麦(Triticum aestivum)、水稻(Oryza sativa)及芦苇(Phragmites australis)等单子叶植物亲缘关系最近;荧光定量RT-PCR分析结果显示,CpPCS基因及CePCS基因受重金属铅锌胁迫上调表达,其中在叶中的表达量显著增加(P<0.05)。通过普陀山苔草植物络合素合酶蛋白基因进行了克隆鉴定,探讨该基因的结构、进化关系及其参与普陀山苔草重金属胁迫的应答模式,为进一步培育重金属污染土壤修复的植物奠定基础。
The hyperaccumulator Carex putuoshan was used as cloning material for the phytochelatin synthase (PCS) gene, and to characterize its expression following heavy metal stress. The results showed that the cDNA of CpPCS was 1 461 bp in length, and it encoded 486 amino acids with a molecular weight of 53.86 kDa and pI value of 6.12. As compared to other plants, the amino acid sequence similarity was approximately 63%. Phylogenetic analysis of PCS proteins from different species showed those of C. putuoshan were most closely related to those of Oryza sativa and Triticum aestivum. Real-time quantitative PCR analysis indicated that CpPCS and CePCS (Carex oshimensis 'Evergold') genes were expressed in the leaves. The CpPCS and CePCS genes were up-regulated in Pb and Zn treatments. Additionally, we investigated CpPCS structure, phylogenetic relationships, and gene expression patterns. Our results indicated that CpPCS might play an important role in heavy metal stress, and could be further utilized in plants.
植物修复技术指利用超富集植物富集土壤中的重金属, 并将根系富集的重金属运输到植物的地上部分, 从而降低土壤中的重金属水平[1]。其对土壤重金属污染修复有着效果好、对环境干扰小、技术成本低、无二次污染和环境美学的兼容性等优点[2]。在超富集植物的研究中, 对抗重金属胁迫的关键基因进行克隆及表达分析, 对筛选与利用超富集植物具有重要意义[1]。植物络合素(phytochelatins, PCs)是细胞响应重金属胁迫而合成的一类巯基多肽化合物, 也被叫做植物螯合肽, PCs不仅响应镉(Cd)的胁迫, 而且能降低其它重金属以及非金属对植物的毒害作用, 提高植物对这些元素的抗性, PCs还能够维持细胞内环境中金属离子浓度的平衡, 它既可以螯合细胞中过量的金属离子, 保护细胞中酶的活性, 同时也可以将这些金属离子保存在细胞体内, 在植物体需要的合适时机释放到细胞环境中[3]。植物络合素合酶(phytochelatin synthases, PCS)是PCs合成途径中的关键酶, 首次在Grill和Zenk[4]的研究中发现, 他们从膀胱麦瓶草(Silenecucublue)细胞中提纯出一种酶, 这种酶能够以GSH为底物而催化合成PCs, 被称为PCs。因而, PCS基因成为植物富集土壤重金属的关键因素之一, 多年来一直受到学者们的重视。自1999年成功克隆拟南芥(Arabidopsis thaliana)、粟酒裂殖酵母(Ascomycetes)和小麦(Triticum aestivum)的AtPCS1、 SpPCS及TaPCS13三个PCS基因以来[5, 6, 7], 已在多种生物中克隆分析了PCS基因, 包括小麦、大蒜(Allium sativum)、狗牙根(Cynodon dactylon)、大豆(Glycine max)、莴苣(Lactuca saliva)、百脉根(Lotusc orniculatus)、豆梨(Pyrus calleryana)、番茄(Lycopersicon esculentum)、印度芥菜(Brassica juncea)、天蓝遏蓝菜(Thlaspi caerulescens)和蜈蚣草(Eremochloa ciliaris)等[8, 9, 10, 11, 12, 13, 14, 15, 16, 17]。将拟南芥AtPCS1基因转入普通烟草后发现, 转基因烟草幼苗的Cd积累量增加了两倍[18]。在烟草中PCs过量表达, 转基因植株在水培条件下对Cd离子的吸收增加了9倍[19]。研究表明, 植物络合素合成酶对于植物富集重金属研究具有重要的意义。
普陀山苔草(Carex putuoshan)是四川农业大学环境学院朱雪梅团队最新发现的一种铅超富集植物, 其繁殖力强, 能生长于大部分生态环境脆弱区, 且生物量大, 是植物修复重金属污染土壤的优质材料。其地上部分对重金属铅的富集量可达2 369.26 mg· kg-1, 远超一般超富集植物标准, 同时, 当锌浓度为125 mg· L-1时, 铅富集达最大[20]。叶片是普陀山苔草地上部分的主体, 分析不同胁迫条件下CpPCS基因在叶中的表达情况, 对于深入理解超富集植物的富集机制具有十分重要的参考价值。因此, 本研究以超富集植物普陀山苔草为研究对象, 以苔草属非超富集观赏性植物金叶苔草(Carex evergold)为对照, 首次尝试从超富集植物— — 普陀山苔草中克隆PCS基因, 并利用实时荧光定量PCR技术研究铅锌复合胁迫下PCS基因的胁迫应答响应情况, 对普陀山苔草的分子应答机制进行初步探索, 进而为普陀山苔草的富集机理及重金属抗性研究提供参考, 并为利用转基因技术开发高效的超富集转基因植物修复系统提供理论依据。
普陀山苔草采集于四川省雅安市汉源县普陀山铅锌矿区。金叶苔草购于花草市场。
将已长出两片以上真叶, 长势良好的普陀山苔草及金叶苔草幼苗移栽至塑料盆(盆口径为13 cm, 高为12 cm)中进行砂培, 每盆栽4株, 每组处理3次重复。整个培养期间, 每7 d浇一次Hoagland完全营养液(100 mL· 盆-1), 每两天浇一次去离子水(100 mL· 盆-1)。对预培养14 d后的苔草进行铅锌处理, 重金属溶液为100 mL· 盆-1。
前期研究发现, 当锌(Zn)浓度为125 mg· L-1、铅(Pb)浓度为600 mg· L-1时, 普陀山苔草富集量和转运系数超过了铅超富集植物的标准[20]。因此, 胁迫的重金属溶液浓度处理设置为0、Pb2+ 600 mg· L-1、Zn2+125 mg· L-1和Pb2+ 600 mg· L-1+ Zn2+125 mg· L-1。分别于胁迫后的0、12、24和36 h对苔草的叶片与根进行取样收集。各处理在每个时间点取3株样为重复。取样时, 叶片用去离子水冲洗3次, 吸干表面水分, 液氮速冻, 放于-70 ℃冰箱保存待用。
1.2.1 RNA提取及反转录 超净工作台、研钵、移液器等试验用具的清洁均按RNA提取的常规方法处理, 参考柱式植物RNAout 3.0 Kit操作手册并稍加改进, 利用电泳和分光光度计检测总RNA样品。取1 μ L苔草总RNA样品, 2%的非变性琼脂糖凝胶电泳, 观察28S rRNA和18S rRNA的亮度比。取1 μ L总RNA、9 μ L 1× TE于比色皿, 混匀, 在RNA程序下测定样品相应值, 样品RNA含量(μ g· mL-1)=40× OD260× n÷ 1 000。采用PrimeScriptTM RT reagent Kit with gDNA Eraser(PerfeCp Real Time)(Takara Code:RR047A)合成cDNA第一链。
1.2.2 两种苔草CpPCS基因cDNA保守区及普陀山苔草全长克隆 植物络合素合酶引物根据GenBank数据库中公布的大蒜(AAO13809.1)、小麦(AF093752.1)、水稻(AF439787.1)、芦苇(AFU06381.1)等氨基酸保守序列, 利用codehop设计简并引物UP和DP。根据上述CpPCS基因的保守片段, 设计3'-RACE引物和5'-RACE引物。序列拼接获得基因全长序列, 以此设计基因特异性引物CpPCS-5'和CpPCS-3', 克隆CpPCS基因的CDS片段。
CpPCS与CePCS基因保守片段的克隆:以合成的cDNA第一链为模板, 用引物UP和DP扩增CpPCS与CePCS基因的保守片段。反应体系(25 μ L):Premix Taq Version 2.0 9.5 μ L, ddH2O 12.5 μ L, 上下游引物各1 μ L, cDNA模板1 μ L 。反应参数分别为:94 ℃ 5 min; 94 ℃ 60 s, 57 ℃ 50 s, 72 ℃ 1 min, 30循环; 72 ℃ 10 min; 94 ℃ 5 min; 94 ℃ 60 s, 60 ℃ 50 s, 72 ℃ 1 min, 30循环; 72 ℃ 10 min。
CpPCS基因3' 端cDNA的克隆:以3' RACE-ready cDNA为模板, 用引物P3R和试剂盒中通用引物扩增CpPCS基因的3'片段。反应体系(25 μ L):Premix Taq Version 2.0 9.5 μ L, ddH2O 12.5 μ L, 上下游引物各1 μ L, L cDNA模板1 μ L。反应参数:94 ℃ 5 min; 94 ℃ 60 s, 50 ℃ 50 s, 72 ℃ 1 min, 30循环; 72 ℃ 10 min。
CpPCS基因5' 端cDNA的克隆:以5' RACE-ready cDNA为模板, 用引物P5R和试剂盒中通用引物扩增CpPCS基因的5'片段。反应体系(25 μ L):Premix Taq Version 2.0 9.5 μ L, ddH2O 12.5 μ L, 上下游引物各1 μ L, cDNA模板1μ L。反应参数:94 ℃ 5 min; 94 ℃ 60 s, 54 ℃ 50 s, 72 ℃ 1 min, 30循环; 72 ℃ 10 min。
CpPCS基因CDS序列的克隆:利用扩增CDS序列的基因特异引物CpPCS-3'和CpPCS-5', 以普陀山苔草cDNA第一链为模板, PrimeSTAR® Max DNA Polymerase扩增CpPCS基因的CDS序列。反应参数:94 ℃ 5 min; 94 ℃ 60 s, 56 ℃ 50 s, 72 ℃ 1 min, 30循环; 72 ℃ 10 min。
各轮PCR扩增所用引物如表1。PCR产物经1.5%琼脂糖凝胶电泳分离, 按TIANgel Maxi Purification Kit操作手册, 稍加改动, 回收目的条带。将PCR产物与pMD19-T载体连接, 连接产物用于制备E. coli化学感受态细胞, 并进行重组质粒的转化、筛选及测序。
1.2.3 CpPCS基因序列分析和系统进化树构建 将得到的普陀山苔草CpPCS基因测序结果进行拼接, 利用Premier 5.0翻译cDNA序列为氨基酸序列, 在NCBI Conserved Domain Search (http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)上搜索保守结构域。 使用MEGA5.0软件, 通过邻接法(neighbor-joining, NJ)构建系统进化树; 利用ExPASy (http://www.expasy.org/tools/)在线软件分析氨基酸序列的基本理化信息; 使用SWISS-MODEL (http://swissmodel.expasy.org/)分析蛋白质三维结构。
1.2.4 CpPCS及CePCS基因表达分析 取胁迫处理后的两种苔草叶100 mg, RNA提取及反转录同1.2.1合成cDNA第一链。利用CpPCS及CePCS基因保守区设计特异性引物PCSqf及PCSqr(表1), 内参基因选用组蛋白基因H3(表1), 采用SYBR染料法进行实时荧光定量PCR。
| 表1 引物序列与预期扩增片段大小 Table 1 Primer sequences and expected sizes of PCR products |
反应参数:95 ℃ 1 min; 95 ℃ 5 s, 54 ℃ 25 s, + Plate Read; GOTO 2, 39 more times; Melt Curve 65.0 to 95.0 ℃, increment 0.5 ℃, 5s, + Plate Read; END。按照2-Δ Δ Ct法计算出待测基因相对表达量[21], 应用SPSS及Origin整理试验数据并作图。
经PCR扩增得到普陀山苔草CpPCS基因与金叶苔草CePCS基因的保守片段, 依此设计RACE引物, PCR扩增得到CpPCS基因两端片段, 进而设计全长引物来扩增CpPCS基因全长, 得到完整的CpPCS基因CDS序列。
利用保守片段引物扩增得到两条441 bp的特异片段(图1A), 序列分析其属于植物络合素合酶蛋白基因, 5' 和3' -RACE分别扩增出282和1 200 bp的片段(图1B, C), 均含两端序列的识别标志, 说明成功得到两端, 序列拼接, 设计全长引物扩增得到1 461 bp的片段(图1D)。利用NCBI进行分析, 发现该序列具有典型的植物络合素合酶蛋白基因特征, 因此命名为CpPCS。
 | 图1 PCR扩增结果Fig. 1 The PCR amplification results 注:M1, Marker D 2 000 bp; M2, Marker D 5 000 bp; 1, CpPCS保守区; 2, CePCS保守区; B, 3'端; C, 5'端; D, CpPCS全长 。Note: M1, Marker D 2 000 bp; M2, Marker D 5 000 bp; 1, CpPCS conserved sequence; 2, CePCS conserved sequence; B, 3' sequence, C, 5' sequence; D, CpPCS full-length sequence . |
普陀山苔草CpPCS基因的开放阅读框由1 461个碱基组成, 编码一个含有486个氨基酸残基的蛋白(图2)。估计的相对分子质量为53.87 kD, 预测的等电点(pI)为5.67~7.77, 预测脂肪系数为87.65, 该蛋白的不稳定系数为47.56, 归类于不稳定蛋白。经NCBI网站BLAST比对, 发现普陀山苔草与玉米(Zea mays, NP_001168641.1)、芦苇(Phragmites australis, AFU06381.1)、大蒜(AAO13809.1)、粉蓝烟草(Nicotiana glauca, ABX10958.1)、苎麻(Boehmeria nivea, AHC98018.1)的PCS氨基酸序列相似性分别为64%、63%、63%、63%和62%。
 | 图2 CpPCS核苷酸序列和其推导的氨基酸序列Fig.2 Nucleotides sequence and predicted amino acids sequence of CpPCS gene注:○, 半胱氨酸残基; ▭, Cys-(Xaa)n-Cys(n=0~5); ⎔, 活性必需氨基酸。Note: ○, cysteine residue; ▭, Cys-(Xaa)n-Cys(n=0~5); ⎔, active essential amino acid. |
CpPCS属于植物络合素亚家族, 与其它物种PCS氨基酸序列存在很高的保守性, 其中N端保守性较高, 该基因响应重金属, 并对植物络合过程起作用。CpPCS有19个半胱氨酸(Cys)残基散布在基因的不同区域上(图2中圆圈表示), 并组合成5个Cys-(Xaa)n-Cys(n=0~5)元件(包括3个Cys-Cys 元件, 图2中方框表示), 其中C371C372XXXC376XXC379是PCS蛋白质典型的重金属离子传感器基因序列[22, 23], 含有3个活性必需氨基酸(Cys56、His162和Asp182)(图2中正六边形表示)。
以CpPCS的氨基酸序列构建物种间的PCS蛋白进化树(图3)可知, 普陀山苔草CpPCS与小麦、水稻(Oryza sativa)及芦苇等单子叶植物亲缘关系最近。在 http://swissmodel.expasy.org上利用Automatic Modelling Mode预测了普陀山苔草 CpPCS蛋白的三维结构, 同时与拟南芥AtPCS1蛋白、蜈蚣草PvPCS蛋白三维结构进行比较发现, 这3种蛋白具有类似的空间结构, 并且可以预测, 它们的功能在植物体中相似(图4)。上述结果表明, 本研究克隆得到的CpPCS基因的氨基酸序列所编码的蛋白为植物络合素合酶。
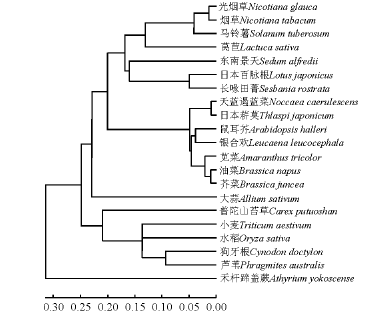 | 图3 不同物种PCS蛋白的系统进化树分析Fig.3 Phylogenetic analysis of PCS proteins from different species |
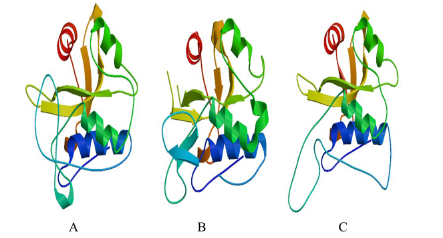 | 图4 蛋白的三维结构预测Fig.4 Ppredicted 3D protein structure注:普陀山苔草CpPCS (A), 印度芥菜BjPCS1 (B), 拟南芥AtPCS (C)Note: Carex putuoshan CpPCS (A), Brassica juncea BjPCS1 (B), Arabidopsis thaliana AtPCS (C) |
除Zn2+胁迫12 h以外, 其它重金属胁迫处理后CpPCS基因在叶中的相对表达量均显著增加(P< 0.05), Pb2+胁迫处理12、24和36 h后, 叶中的相对表达量提高到不胁迫时的2.50、2.19、3.45倍。Zn2+胁迫处理24和36 h后, 叶中的相对表达量提高到不胁迫的1.94倍和4.53倍, Pb2++Zn2+复合胁迫处理12、24和36 h后, 叶中的相对表达量提高到不胁迫时的2.85、2.29、12.58倍(图5)。CePCS基因在金叶苔草叶中表达, 重金属胁迫下该基因相对表达量也显著增加, Pb2+胁迫处理12、24、36 h后, 叶中提高到不胁迫时的1.41、1.35、1.77倍, Zn2+胁迫处理12、24、36 h后, 叶中提高到不胁迫时的1.01、1.92、7.16倍, Pb2++Zn2+复合胁迫处理12、24、36 h后, 叶中提高到不胁迫时的1.35、2.36、9.175倍(图5)。以上结果说明不同重金属对其相对表达量的诱导能力为Pb2++Zn2+> Zn2+> Pb2+。
 | 图5 重金属胁迫下普陀山苔草CpPCS和CePCS基因在叶中的相对表达量Fig.5 Expression of CpPCS and CePCS gene in leaves of Carex putuoshan under heavy metal stress注:不同小字母表示各处理间差异显著 (P< 0.05)。Note: Different lower case letters indicate significant difference among different treatments at the 0.05 level. |
本研究克隆了普陀山苔草CpPCS基因, PCS蛋白氨基酸长度为465~506 aa, 预测等电点为5.67~7.77; PCS蛋白主要定位于细胞核中; 植物中PCS蛋白大多为不稳定蛋白, PCS蛋白的二级结构由α 螺旋、无规则卷曲以及延伸链等元件组成, 空间构架极其相似。本研究得到的CpPCS蛋白为不稳定蛋白, 具有亲水性并且有两个跨膜区域, 具有一个典型的植物络合素合成酶功能域。BLAST 比对发现, 该基因与其它植物PCS基因氨基酸序列同源性达60%以上, 其中与小麦ZmPCS基因同源性达64%, 表明该基因在进化过程中具有较高的保守性; 氨基酸序列比较发现PCS蛋白的N端在不同物种之间具有高度的保守性, 同源性高达40%~50%, 而C端却是多变的[24], Cys残基的位置和排列方式(特别是 N 末端区域)对植物络合素合酶蛋白活性和镉耐受能力非常重要。这种Cys的排列类似于MT中Cys的排列方式[25, 26]。
相邻的Cys元件是螯合重金属离子的重要功能元件, 有研究表明[27], 将酵母PCS基因中Cys173突变为Ala173后, 其结合Cd离子的活性完全消失, 并且降低了PCs的合成效率。C358C359XXXC363XXC366基序是植物络合素合酶的重金属离子传感器[28], 在百脉根LjPCS1和豆梨PcPCS1蛋白中表现为C368C369RETC373MKC376和C369C370QETC374VKC377形式, 而苎麻BnPCS1蛋白中表现与豆梨完全相同[29], 而普陀山苔草CpPCS蛋白中表现为C371C372KETC376VKC379。推测该基序在普陀山苔草中同样行使重金属离子传感器功能, 响应重金属胁迫的重要功能元件。通过三维结构预测可以看出, 普陀山苔草PCS与拟南芥和印度芥菜具有相似的空间构架, 它从侧面印证PCS蛋白是普陀山苔草的重金属富集机制的重要组成部分。
叶片是普陀山苔草地上部分的主体, 同时也是普陀山苔草PCS基因的主要合成部位。分析不同胁迫条件下CpPCS基因在叶中的表达情况, 对于深入理解超富集植物的富集机制具有十分重要的参考价值。qRT-PCR分析表明, CpPCS基因受重金属铅锌胁迫诱导上调表达, 其中在叶中的表达量显著增加, 这与豆梨PcPCS1[13]、萝卜(Raphanus sativus)RsPCS[30]基因的表达特征一致。这可能是普陀山苔草地上部分富集量高的原因之一。
单一铅锌胁迫下, CpPCS及CePCS的表达量大部分随着时间变化呈现先升高后略微降低再升高的趋势, 这说明铅锌处理均会诱导CpPCS及CePCS的表达, 且锌处理下表达量稍高, 这与李慧等[13]及姜倩倩等[31]的研究相符合。而在铅锌符合复合胁迫下, 两种苔草PCS基因均高表达, 特别是普陀山苔草叶片CpPCS基因表达量达到空白对照组的12倍, 是相同处理下金叶苔草的8倍。从一方面来说, 在铅锌胁迫环境下, 铅锌均可能是普陀山苔草中CpPCS的诱导因素之一, 复合胁迫下铅锌的诱导产生协同作用, 使普陀山苔草CpPCS大量表达, 刺激了植物络合素高效快速的合成, 才使得普陀山苔草能在高浓度的铅胁迫下有很强的耐性, 这可能是普陀山苔草富集作用的原因之一。从另一方面来说, 则可能是普陀山苔草CpPCS大量表达, 刺激了植物络合素高效快速的合成, 使得虽然普陀山苔草不富集锌却能耐受锌的毒害。这说明CpPCS是铅锌胁迫下重要的解毒基因与富集基因。
(责任编辑 苟燕妮)
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|