


{kind=link}
{kind=link}
{kind=link}
{kind=link}
南方红壤丘陵区3种森林群落内主要草本植物种群生态位特征
[潘高1, 2  , 张合平
, 张合平1 , 潘登1, 2 ]
, 张合平, 潘登|
|


第一作者:潘高(1991-),男,湖南岳阳人,在读硕士生,主要从事森林生态学研究。E-mail:windypg@126.com
生态位是解释自然群落中种间共存与竞争机制的基本理论,对了解植物群落构建及其与环境因子的耦合关系具有重要意义。本研究以南方红壤丘陵区3种森林群落内主要草本植物为研究对象,在群落调查的基础上,运用Levins生态位宽度、Pianka生态位重叠指数和生态位相似性公式,分析了针叶林、阔叶林和针阔混交林下主要草本物种的生态位宽度以及物种间的生态位重叠与生态位相似性,并从生态位角度探讨了不同森林群落内草本种群的生长状况。结果表明,3种森林群落内草本种群的生态位宽度平均值从大到小依次为针叶林(0.572)>针阔混交林(0.508)>阔叶林(0.424),且针叶林中崖爬藤( Tetrastigma formosanum)、淡竹叶( Lophatherum gracile)、鳞毛蕨( Dryopteris sp.)以及六月雪( Serissa japonica)等草本植物的生态位宽度均大于针阔混交林和阔叶林的;生态位重叠度与生态位宽度之间不是绝对的正相关关系,生态位宽度小的物种与其他种群也可能发生较大的生态位重叠,甚至出现完全重叠的现象;针叶林和阔叶林内主要草本植物的生态位宽度与其重要值呈显著正相关( P<0.01),相关系数分别为0.758和0.918;3种森林群落内草本种群的生态位宽度与其重要值变异系数均呈显著负相关( P<0.01),相关系数分别为-0.996、-0.967和-0.963。在现阶段植被演替过程中,针叶林和针阔混交林下草本种群的重叠程度较高,环境资源存在着高度的空间异质性,群落结构仍处于不稳定状态。
The niche theory has been successfully used to explain competition mechanisms among the coexisting species in natural communities, which is significant for understanding of the coupling between plant community construction and environmental factors. The niche characteristics of herb populations were examined within coniferous forest(CF), broad-leaved forest(BF) and mixed broadleaf conifer forest(MF) in hilly red soil region of southern China, using Levins niche breadth, Pianka index of niche overlap and niche similarity formula, from the perspective of the growth status in different forest communities in herbage species. Results showed, the mean niche breadth of all herbage species in the studied forest communities was in order CF(0.572)>MF(0.508)>BF(0.424). The niche breadth of Tetrastigma formosanum, Lophatherum gracile, Dryopteris sp., Serissa japonica and other herbaceous plants in CF were higher than that in MF and BF. The niche overlap did not always correlate to niche breadth. Larger niche overlap or even completely overlapping might occurr between the species with narrow niche breadth. There was a significant positive correlation between the niche breadth and the important value of the main herbaceous plants in CF and BF( P<0.01),with the correlation coefficients of 0.758 and 0.918, respectively. There was a significant negative correlation between the niche breadth and the important value of the coefficient of variation of herbage species in three forest communities( P<0.01), with the correlation coefficients of -0.996, -0.967 and -0.963, respectively. Our study suggested the CF and MF were still in an instability state at present stage of community succession due to high niche overlap among herb populations and high spatial heterogeneity of environmental resources in study region.
生态位(Niche)作为进行自然群落中种间共存与竞争机制研究的基础, 通过定量地研究植物种、植物群落及其与环境之间的耦合关系, 进一步揭示植物种群利用自然资源的能力、占据生态空间的范围和相对地位, 对森林生态系统的恢复与重建具有重要的理论和现实意义[1, 2, 3]。关于生态位的研究, 国内外已经做了大量工作[4, 5, 6, 7, 8]。国内以往的研究大多集中在研究森林群落内乔木与灌木优势种群种间关系[9]、群落结构[10]、物种多样性保育[11]及群落演替与恢复[12], 而关于草本种群生态位的研究较少[13]。那么, 在同一森林群落内不同草本种群是如何相互适应?草本植物对空间资源的利用途径以及在不同森林群落内是否存在一些共同特征?这两个科学问题有待相关的研究案例来探讨与论证。
南方红壤丘陵区被认为是我国最脆弱的生态系统之一, 区域自然地理特征和长期的人为干扰共同导致植被稀疏、基岩裸露、土地生产力低下, 使其成为我国水土流失程度仅次于黄土高原的地区[14, 15]。长期以来, 该区域开展了大量的人工林营造和植被恢复工作, 但森林覆盖率的增幅与水土流失面积的减幅极不相称, 这主要是由于林下灌木与草本植物覆盖不足, 致使林地土壤结构恶化, 植被恢复十分困难[16, 17]。为此, 本研究以南方红壤丘陵区3种森林群落中主要草本植物为研究对象, 运用Levins生态位宽度、Pianka生态位重叠指数和生态位相似性公式对针叶林、阔叶林和针阔混交林下草本物种的生态位宽度以及物种间的生态位重叠与相似性进行定量研究, 旨在了解主要植物种群在群落中的功能和地位, 探讨物种对环境资源的生态适应性, 为进一步研究中亚热带森林生物多样性保育及其恢复机制提供参考。
研究区位于青羊湖国家森林公园, 地处湖南省长沙市宁乡县西部、沩水上游, 地理坐标为28° 07'-28° 14' N、111° 54'-112° 05' E。北及东北与桃江县相接, 东南方向毗邻黄材镇, 西与安化县接壤, 公园总面积1 129.06 hm2。研究区为大陆性季风气候, 属亚热带湿润气候区, 冬寒夏热, 春秋短促, 冬夏绵长, 年平均气温16.2~17.6 ℃, 年平均日照1 737.6 h, 无霜期273 d, 年平均降水量1 358.3~1 471.3 mm。该区为典型的低山丘陵地貌, 海拔高度在50~326 m, 坡度平缓, 一般在45° 以下, 土壤以板页岩发育而成的红壤为主, 其次是黄红壤、黄棕壤, 土壤质地以壤粘土为主。植物群落以马尾松(Pinus massoniana)、杉木(Cunninghamia lanceolata)为主的针叶林, 以青冈(Cyclobalanopsis glauca)、南酸枣(Choerospondi asaxillaris)、枫香(Liquidambar formosana)为主的阔叶林和以马尾松(Pinus massoniana)、杉木、南酸枣、枫香、青冈为主的针阔混交林较为典型; 灌木主要有山茶(Camellia japonica)、檵木(Loropetalum chinensis)、盐肤木(Rhus chinensis)、黄檀(Dalbergia hupeana)等; 草本植物主要有苔草(Carex schmidtii)、狗脊蕨(Woodwardia japonica)、鳞毛蕨(Dryopteris sp.)、淡竹叶(Lophatherum gracile)等。
2013年7-8月对青羊湖国家森林公园进行了全面踏查。本研究采用典型样地调查法, 在充分考虑海拔、坡位、坡向、坡度等立地条件的基础上, 选取有代表性的群落类型, 共设置15块调查样地, 样地基本情况如表1。3种群落类型均分别设置5块20 m× 30 m的标准地进行群落调查, 对样地内所有高度≥ 3 m的乔木进行每木检尺, 并记录种类、个体数、胸径、高度、冠幅(东西、南北)等信息; 在每块样地四角及中点设置5个5 m× 5 m的灌木样方调查高度≥ 1 m的灌木, 同时在每个灌木样方内随机设置1个1 m× 1 m的草本小样方调查草本植物多样性, 共调查灌木样方75个、草本样方75个, 并记录所有灌木(包括木质藤本及乔木更新幼苗、幼树)、草本(包括草质藤本)的植物名称、株数、高度、盖度等指标。
重要值为描述草本植物在其所在层片中的功能地位, 反映其分布格局, 采用重要值(Important Value, IV)作为测度植物种群在群落中的优势度指标。根据样地调查记录的草本植物种类, 计算各样地内不同物种的重要值, 然后统计出3种森林群落内主要草本种群的重要值, 取其平均值, 选择重要值> 1的物种作为本研究进行生态位研究的种群。其计算公式如下[18]:
IV(重要值)=相对密度+相对频度+相对盖度。
相对密度指样地内某一物种的个体数占全部物种个体数的百分比, 计算公式:
相对密度=某物种的个体数/所有物种的个体数× 100%;
相对频度指某一物种在全部样方中的频度与全部物种频度总和之比, 计算公式:
相对频度=某物种的频度值/所有物种的频度值× 100%;
相对盖度指样地内某一物种的分盖度占所有分盖度之和的百分比, 计算公式:
相对盖度=某物种的分盖度/所有物种的分盖度× 100%;
生态位宽度(Bi)采用Levins提出、后经Corwell修改的计算公式[19]:
Bi=1/r
式中, Bi为物种i的生态位宽度; Pij为物种i在资源j上的重要值占该种在所有资源上重要值的比例; r为资源位总位数。
生态位重叠度(Oik)采用Pianka计算公式[20]:
Oik=
式中, Oik为物种i与物种k的生态位重叠指数; Pij和Pkj分别为物种i和物种k在资源j上的重要值占该种在所有资源上重要值的比例; 其他符号含义同上。
生态位相似性(Cik)多认为是不同种群利用资源的相似程度, 计算公式[21]:
Cik=1-
式中, Cik为物种i与物种k的生态位相似性比例值; 且Cik=Cki; Pij和Pkj分别为物种i和物种k在资源j上的重要值百分率。
采用SPSS 19.0对各森林群落内草本植物的生态位宽度、重要值及其重要值变异系数进行Pearson相关性分析, 在Excel 2003中分别计算物种的重要值、生态位宽度、生态位重叠度和生态位相似性等指标; 并利用SigmaPlot 12.5软件绘制相关分析图形。
针叶林中重要值最大的分别为淡竹叶(319.17)、鳞毛蕨(223.08)、芒萁(218.23)和檵木(102.92), 其次为狗脊蕨(83.19)、崖爬藤(80.61)、崖豆藤(62.15)和苔草(42.46)等, 表明淡竹叶、鳞毛蕨、芒萁和檵木为南方红壤丘陵区针叶林群落内草本层的优势物种(表2)。在阔叶林中重要值最大的依次为狗脊蕨(279.68)、鳞毛蕨(275.10)、苔草(235.73)和淡竹叶(126.99), 分别占整个阔叶林群落重要值之和的23.15%、22.77%、19.51%和10.51%, 是该群落的主要优势种群, 其他还有绣线菊(47.38)、崖爬藤(36.69)、菝葜(32.10)和忍冬(31.09)等(表3)。针阔混交林中鳞毛蕨(308.01)、狗脊蕨(266.44)和淡竹叶(113.96)的重要值最大, 在草本层中占有重要地位, 是该群落的优势种, 其次为苔草(77.59)、乌敛莓(58.22)、六月雪(49.52)以及崖豆藤(48.59)、菝葜(47.99)和海金沙(43.46)等(表4)。
| 表2 针叶林群落主要草本植物的重要值 Table 2 Importance value (IV) of main herb populations in coniferous forest |
| 表3 阔叶林群落主要草本植物的重要值 Table 3 Importance value (IV) of main herb populations in broad-leaf forest |
| 表4 针阔混交林群落主要草本植物的重要值 Table 4 Importance value (IV) of main herb populations in mixed broadleaf-conifer forest |
在针叶林中, 崖爬藤、淡竹叶、鳞毛蕨和芒萁生态位宽度值较大, 分别为0.878、0.826、0.783和0.690, 表明崖爬藤等4种草本植物分布范围广, 同时具有较强的环境资源利用能力, 是针叶林群落的优势种。阔叶林中生态位宽度值从大到小依次为苔草> 狗脊蕨> 鳞毛蕨, 分别为0.823、0.750和0.733, 其他大多数物种的生态位宽度较小, 尤其是崖爬藤、崖豆藤和班茅的生态位宽度值均为0.200, 说明这几个物种所占据生态空间的范围较窄, 也反映出阔叶林下草本群落结构的单调性(表5)。在针阔混交林群落的13种草本植物中, 苔草的生态位宽度值最大, 为0.881, 其次是鳞毛蕨(0.781), 表明他们具有较强的生态适应力; 乌敛莓、薯蓣、青冈和忍冬的生态位宽度值较小, 分别为0.399、0.385、0.385和0.200, 且在实际调查中也发现, 这4个物种在针阔混交林的空间分布异质性较高, 反映出物种自身的生物学和生态学特性导致其适宜的生境减少。针叶林主要草本植物种群的生态位宽度值主要集中在0.3~0.7及> 0.7区间的生态位宽度分布较为平均(图1), 对资源环境的利用程度较为均衡。在3种森林群落内, 针阔混交林下草本物种的生态位宽度分布更为平均, 表明该群落的草本植物种群对群落内部环境中绝大多数资源的利用更均衡。
淡竹叶、鳞毛蕨、六月雪、崖豆藤、狗脊蕨、苔草和海金沙7种植物在3种森林群落内均有分布, 说明这几个物种在当地的生态适应力相对其他草本植物强, 对环境资源的利用更为宽泛。同一植物在不同群落中的生态位宽度差异较大, 如崖爬藤在针叶林群落的生态位宽度最大, 为0.878, 在阔叶林中生态位宽度仅为0.200, 在针阔混交林群落中却未见分布; 苔草在阔叶林、针阔混交林群落中生态位宽度最大, 分别为0.823和0.881, 而在针叶林中生态位宽度居中。
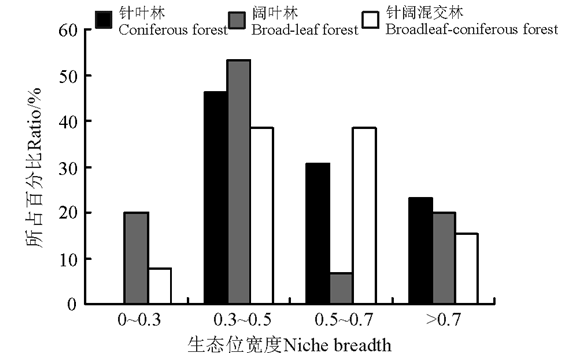 | 图1 3种森林群落内主要草本种群生态位宽度的物种百分比Fig.1 The ratio of niche breadth of main herb populations in three forest types |
| 表5 针叶林、阔叶林和针阔混交林群落主要草本种群的生态位宽度 Table 5 Niche breadth of main herb populations in coniferous, broad-leaf and broadleaf-conifer forest |
3种森林群落草本植物种群的生态位宽度平均值从大到小依次为针叶林> 针阔混交林> 阔叶林, 说明针叶林和针阔混交林群落的草本物种对资源的利用率较高, 更有利于林下植被的更新和演替。针叶林内主要草本种群的生态位宽度均大于针阔混交林、阔叶林群落, 说明在南方红壤丘陵区针叶林下草本群落较针阔混交林和阔叶林具有生态幅度广、资源竞争能力强的特征, 能更充分地利用生境中的绝大数资源。
在3种森林群落内草本植物种群的生态位重叠度差异很大, 针叶林、阔叶林和针阔混交林下草本种群生态位重叠度在0~0.3的物种百分率分别为16.48%、43.33%和27.47%, 生态位重叠度> 0.7的物种百分率依次为43.96%、32.50%和31.87%, 生态位重叠平均值趋势表现为针叶林(0.609)> 针阔混交林(0.543)> 阔叶林(0.446), 与生态位宽度平均值的结果一致(图2)。这表明在南方红壤丘陵区针叶林群落的物种较针阔混交林、阔叶林更加丰富, 对环境资源的利用也更为宽泛, 同时由于针叶林下草本群落生态位宽度平均值最大, 草本植物种群实现了自身优化, 相应也使得物种间的竞争更为激烈, 在物种不断适应环境和物种间相互竞争的共同作用下, 群落内部生境的资源可以得到最大限度的利用。
 | 图2 3种森林群落内主要草本种群生态位重叠度的物种百分比Fig.2 The ratio of niche overlapping of main herb populations in three forest types |
阔叶林群落草本植物种群的生态位重叠度平均值较小, 仅为0.446, 这主要是该群落草本植物生态位宽度较窄, 林下草本物种多样性较少, 导致各植物种生态位分化明显, 也可能是由于物种自身生物生态学特性不同、共同具有的资源位少以及对生境条件的要求不同。
生态位宽度与生态位重叠度之间存在一定的相关性, 生态位宽度值较大的物种与其他物种的生态位重叠度通常也较高。如:鳞毛蕨在3种森林群落内都具有较大的生态位宽度值, 在针叶林中, 鳞毛蕨和崖爬藤的生态位重叠度为0.877, 与淡竹叶的生态位重叠度为0.841(表6); 在阔叶林中与淡竹叶的生态位重叠度为0.886(表7); 在针阔混交林中, 鳞毛蕨与苔草、黄檀的生态位重叠度分别为0.875和0.822(表8)。这是因为鳞毛蕨在群落生境中处于优势地位, 具有较强的占据生态空间和资源竞争能力, 从而与其他物种的生态位重叠的机会也更高。但有些物种生态位宽度较小却与其他物种的生态位重叠度较高, 如:针叶林群落中海金沙与土牛膝的生态位重叠度为0.996; 在阔叶林中, 山茶和青冈的生态位重叠度高达0.998。这可能是为了在资源或活动空间受到限制的生境中生存, 生态位窄的物种与其他种群的生物生态学特性及对资源环境的要求趋于基本一致, 在进行激烈种间竞争后的结果。
| 表6 针叶林群落下主要草本种群生态位重叠值 Table 6 Niche overlap of main herb populations in coniferous forest |
| 表7 阔叶林群落下主要草本种群生态位重叠值 Table 7 Niche overlap of main herb populations in broad-leaf forest |
| 表8 针阔混交林群落下主要草本种群生态位重叠值 Table 8 Niche overlap of main herb populations inmixed broadleaf-conifer forest |
在针叶林草本群落13个主要种群组成的78个种对中, 生态位相似性比例值为0的有4对, 占总数的5.13%, 生态位相似性比例值在0.5以上的物种共35对, 占总数的44.87%, 说明多数物种利用环境资源的相似程度较高, 生态位分化不明显; 另外, 生态位宽度较小的物种狗脊蕨与薯蓣、海金沙与土牛膝之间的生态位相似性比例值超过0.895, 这表明生态位宽度与生态位相似性之间并不存在绝对的正相关关系(表9)。
| 表9 针叶林群落下主要草本种群生态位相似性 Table 9 Niche similarity of main herb populations in coniferous forest |
在阔叶林群落下, 生态位相似性比例值为0的物种共27对, 占总数的25.71%, 其中在0.1~0.4的有37对, 占总数的35.24%, 说明这些物种对环境资源的要求差异性较大; 生态位相似性比例值在0.5以上的共28对, 占总数的26.67%, 同时, 山茶与青冈以及崖爬藤与班茅的生态位相似性比例值高达0.960和1.000, 再次说明了生态位宽度小的物种间也可能出现较大的生态位相似性, 这主要与物种自身的生物生态学特性以及对生境因子的要求相似程度有关(表10)。
在针阔混交林下, 13个草本种群间的生态位相似性比例值差异显著, 其中在0.1~0.4的共28对, 占总数的35.90%, 生态位相似性比例值为0的物种有6对, 占总数的7.69%, 另外, 还有23个种对的生态位相似性比例值在0.5以上, 占总数的29.49%, 这表明在针阔混交林群落内大多数草本物种间的生态位相似性程度较低, 生态位分化明显, 潜在的种间竞争并不激烈(表11)。
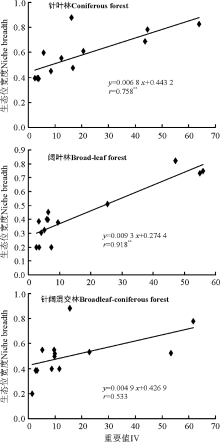
在3种森林群落内, 针叶林和阔叶林群落主要草本种群生态位宽度与其重要值呈典型正相关关系, 其相关性均达到显著水平(P< 0.01), 相关系数分别为0.758和0.918; 而针阔混交林下生态位宽度与重要值的相关性尚未达到显著水平(P> 0.05), 呈微弱的正相关关系(图3)。各森林群落内草本植物种群的生态位宽度与其重要值变异系数均呈典型的负相关关系, 相关系数分别为-0.996、-0.967和-0.963, 且相关性均达到显著水平(P< 0.01), 这表明各种群在样地内的重要值变异系数越大, 其生态位宽度反而越小(图4)。
 | 图3 3种森林群落内主要草本种群生态位宽度与重要值的相关性注:* * , 表明差异极显著(P< 0.05)。图4同。Fig.3 Correlation between the niche breadth and the IV of main herb populations in three forest typesNote: * * , indicate significant different at 0.01 level. The same in Fig.4. |
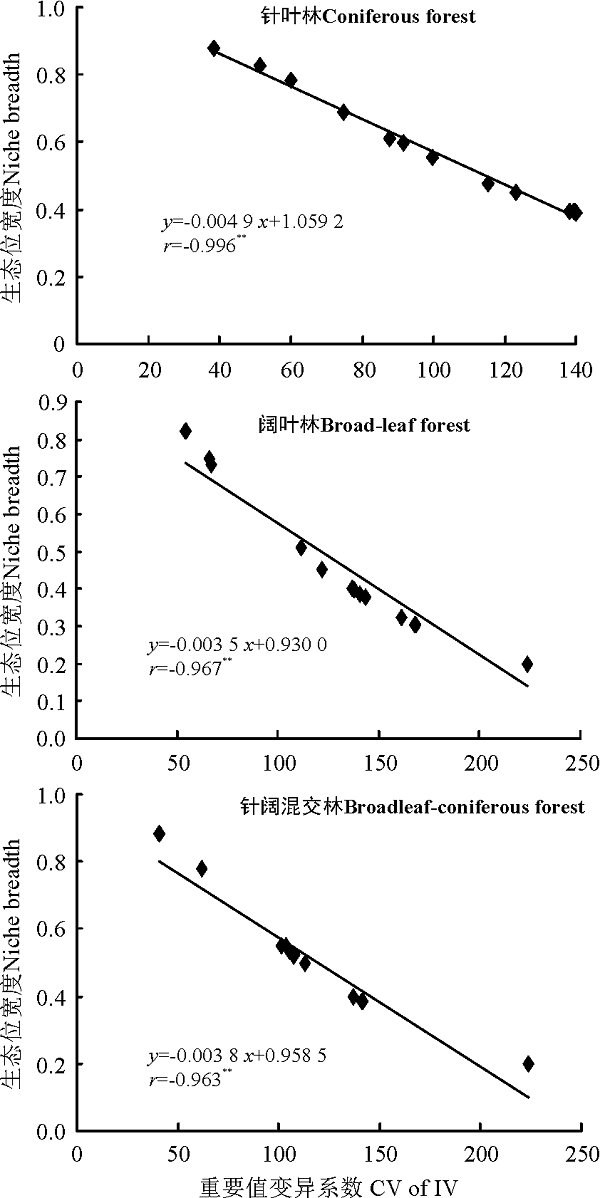 | 图4 3种森林群落内主要草本种群生态位宽度与重要值变异系数的相关性Fig.4 Correlation between the niche breadth and coefficient of variation the IV of main herb populations in three forest types |
生态位反映了自然群落中种群对资源的利用程度, 也表征了他们对环境的生态适应力、生态幅度大小, 是物种适应群落生境内部各生态因子的生存策略和机制[22, 23]。本研究中, 淡竹叶、鳞毛蕨、崖豆藤等7种植物在3种森林群落内均有分布, 但生态位宽度并不一致, 表明不同林分类型下草本群落的广生态幅物种存在较大差异, 这不仅与其自身的竞争能力有关, 也与物种间生物生态学特性和生境内部环境因子的分布状况有关。3种森林群落内草本物种的生态位宽度平均值从大到小依次为针叶林> 针阔混交林> 阔叶林, 且主要草本种群的生态位宽度也表现为针叶林> 针阔混交林> 阔叶林群落。本研究表明, 在南方红壤丘陵区现阶段的植被恢复(演替)进程中, 针叶林下的草本群落对资源环境的利用尺度更大、竞争能力更强, 更有利于林下植被的更新和演替。此外, 还与针叶林内郁闭度低、光照充足以及良好的通风条件等密不可分, 是物种与群落内各种生态因子长期综合作用的结果。
生态位重叠既表征了物种对环境资源利用状况的相似性, 同时也体现了种群间所占据空间位置的交错程度[24]。生态位重叠程度高, 种间竞争强度越大, 越有利于群落演替的发生; 反之, 生态位重叠度小, 种间竞争强度也越小, 群落处于相对稳定的状态。如:针叶林、针阔混交林群落生态位重叠度0.5~0.7及> 0.7所占的比例分别为65.94和61.54, 这说明该森林群落内物种间在共享资源的同时存在着激烈的竞争, 显示现阶段针叶林和针阔混交林群落结构较不稳定。本研究中的一些草本植物, 如针叶林下海金沙与土牛膝, 山茶与青冈以及绣线菊与蛇葡萄在阔叶林群落中的生态位宽度较小, 而生态位重叠度均在0.960以上。这可能与物种分布的不连续性、资源空间异质性高以及小生境的多样性与复杂性有关, 导致某些植物以斑块的形式聚集分布在局部适宜的群落生境中, 而在适宜生境之外的空间分布较少, 以致出现这些物种在整个群落中生态位宽度较小, 却有着较高生态位重叠度的现象。
重要值和生态位宽度均可用来衡量植物种群在群落中的地位和作用, 但两者的意义完全不同, 前者反映的是植物种在群落中的优势度, 后者则表示不同物种对环境资源的利用状况及生态适应幅度[25]。本研究表明, 针叶林和阔叶林群落主要草本物种生态位宽度与其重要值呈典型正相关, 且相关性极显著(P< 0.01), 这与汪建华等[26]在金佛山对植物种群生态位的研究结果相一致。但由于重要值大小与所占据资源位的数量之间没有必然的关联, 两者之间并不总是具有明显的一致性, 即物种重要值高其生态位宽度不一定大。针阔混交林下生态位宽度与重要值虽然也呈正相关关系, 但其相关性并不显著, 如狗脊蕨和黄檀在相同的资源位中出现, 狗脊蕨的重要值是黄檀的10.12倍, 而生态位宽度仅为后者的0.96倍。这是因为狗脊蕨在大多数资源位中均为优势物种, 重要值之和约占总和的1/4, 其重要值变异系数(107.95)大于黄檀的重要值变异系数(103.52)。因此, 物种的生态位宽度与其重要值之和以及在资源位中分布的均匀程度有着密切的关系。另外, 本研究将物种重要值变异系数与生态位宽度进行相关性分析发现, 3种森林群落内草本种群的生态位宽度与其重要值变异系数均呈典型的负相关关系, 且相关性极显著(P< 0.01), 即物种在样地内的重要值变异系数越大, 其生态位宽度反而越小, 这与以往的研究结果一致[6]。物种生态幅度广并不意味着具有很强的资源利用能力。当物种在各资源位的重要值分布出现悬殊时, 重要值低则表明物种对该资源位的利用率不高, 可能出现分布范围广、生态位宽度不大的情形, 如针叶林群落内狗脊蕨占据了3/5的资源位, 重要值变异系数高达115.17, 生态位宽度值为0.477, 居第8位。
南方红壤丘陵区是一个开放的生态系统, 自然的风力、水力以及动物和人为干扰为植物的侵入提供了载体和媒介, 而其本身风化程度高、地形破碎、土层浅薄等恶劣的初始条件, 又对侵入的植物进行了严格的筛选[27]。只有生态位宽度大、对资源的利用能力和对生态因子适应能力强的先锋种群才能成功定居。而崖爬藤、淡竹叶、鳞毛蕨和六月雪等草本植物正是凭借对环境要求不高以及生长力较强的生态学特征成为针叶林、阔叶林和针阔混交林群落的先锋种群乃至优势种群。随着演替的推进, 这些物种所占据生态空间的范围逐渐增大, 并形成具有较高聚集度的斑块状分布格局。群落的发展与演替既是物种对环境的适应过程, 也是环境对物种的自然选择过程[28]。在种间竞争和资源环境压力的双重作用下, 生态位高度重叠的两个物种趋向于相互适应、协同进化, 并在物种形态、生理以及对生态因子等方面有互补性要求, 从而达到物种与环境的和谐统一。因此, 这也可能是南方红壤丘陵区群落生境中许多生态位重叠的物种能够共存, 且处于相对稳定状态的一个原因。
本研究对南方红壤丘陵区3种森林群落内主要草本植物种群的生态位特征进行了分析, 研究发现:1)在现阶段植被演替过程中, 南方红壤丘陵区3种森林群落内主要草本种群间存在着普遍的生态位重叠, 针叶林和针阔混交林下草本种群的重叠程度较高, 物种间对环境资源存在着激烈竞争和高度的空间异质性, 群落结构仍处于不稳定状态。2)物种的生态位宽度与其重要值之间虽然存在一定的相关性, 但由于物种重要值大小与其所占据资源位的数量之间没有必然的关联, 两者之间并不总是具有明显的一致性。因此, 物种重要值高其生态位宽度不一定大。3)物种的生态幅度广并不意味着具有很强的资源利用能力。当物种在各资源位的重要值分布出现悬殊时, 重要值低则表明物种对该资源位的利用不充分, 也可能出现分布范围广、生态位宽度不大的情况。
致谢:中南林业科技大学王光军教授对本论文撰写提出宝贵意见, 美国伊利诺伊州州长州立大学谌小勇教授在英文摘要修改过程中给予帮助, 在此表示感谢!
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|